细胞生物学 第九章 细胞核与染色质
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18 __ __ ___ __
/\ \/\ \ /\_ \ /\ \
\ \ `\\ \ __ __ ___\//\ \ __ __ __ ____ __ ___ \_\ \
\ \ , ` \/\ \/\ \ /'___\\ \ \ /'__`\/\ \/\ \ /',__\ /'__`\ /' _ `\ /'_` \
\ \ \`\ \ \ \_\ \/\ \__/ \_\ \_/\ __/\ \ \_\ \/\__, `\ /\ \L\.\_/\ \/\ \/\ \L\ \
\ \_\ \_\ \____/\ \____\/\____\ \____\\ \____/\/\____/ \ \__/.\_\ \_\ \_\ \___,_\
\/_/\/_/\/___/ \/____/\/____/\/____/ \/___/ \/___/ \/__/\/_/\/_/\/_/\/__,_ /
____ __ __
/\ _`\ /\ \ /\ \__ __
\ \ \/\_\\ \ \___ _ __ ___ ___ ___ __ \ \ ,_\/\_\ ___
\ \ \/_/_\ \ _ `\/\`'__\/ __`\ /' __` __`\ /'__`\ \ \ \/\/\ \ /' _ `\
\ \ \L\ \\ \ \ \ \ \ \//\ \L\ \/\ \/\ \/\ \/\ \L\.\_\ \ \_\ \ \/\ \/\ \
\ \____/ \ \_\ \_\ \_\\ \____/\ \_\ \_\ \_\ \__/.\_\\ \__\\ \_\ \_\ \_\
\/___/ \/_/\/_/\/_/ \/___/ \/_/\/_/\/_/\/__/\/_/ \/__/ \/_/\/_/\/_/
细胞核(nucleus): 细胞核是真核细胞内最大、最重要的细胞器。细胞核主要由核被膜、核纤层、染色质、核仁及核体组成, 是遗传信息的储存场所, 是遗传与代谢的调控中心。除高等植物韧皮部成熟的筛管和哺乳动物成熟的红细胞以外其他真核细胞都有细胞核。
核被膜
核被膜(nuclear envelope): 位于细胞核的最外层, 由两层单位膜所组成, 是细胞核与细胞质之间的界膜。外核膜与内核膜相连, 表面附有大量的核糖体颗粒。主要结构为: 双层核膜、核孔复合体、核纤层。核被膜能够控制物质进出。
功能:
- 核被膜构成了核、质之间的天然选择性屏障, 将细胞分成核与质两大结构与功能区域, 使得DNA复制、RNA转录与加工在核内进行, 而蛋白质翻译则局限在细胞质中。避免了核质间彼此相互干扰, 使细胞的生命活动秩序更加井然, 同时还能保护核内的DNA分子免受损伤;
- 核被膜调控细胞核内外的物质交换和信息交流。核被膜并不是完全封闭的, 核质之间进行着频繁的物质交换与信息交流。这些物质交换与信息交流主要是通过核被膜上的核孔复合体进行的;
- 染色质和酶分子的支架和固着部位。
核膜
核膜结构
两层平行的单位膜构成, 两层膜间为相距20~40nm的空间, 称为核周间隙或者核周池。
特点:
- 外核膜: 通常附有核糖体颗粒, 且常常与糙面内质网(rER)相连, 使核周隙与内质网腔彼此相通。
- 内核膜: 表面光滑, 无核糖体附着, 但内表面附着有致密的核纤层和特有蛋白质成分。核孔(nuclear pore): 双层核膜互相平行但并不连续, 内、外核膜常常在某些部位相互融合形成环状开口, 称为核孔。
核膜的崩解与组装
真核细胞有丝分裂时, 核被膜于前期解体, 末期重现, 进行规律性的解体与重建。
- 有丝分裂前期: 核被膜非随机、有区域特异性的解体, 形成单层膜泡, 核孔复合体消失, 核纤层去组装。
- 有丝分裂末期: 核被膜围绕染色体重建, 旧核膜与膜泡参与这一过程。首先附着于染色体表面, 并相互融合形成双层核膜, 同时膜上的某些功能区域相互融合, 与蛋白质组装形成核孔复合体。
- 核被膜的解体与重建受到细胞促进成熟因子(MPF)的调控, 与核纤层蛋白、核孔复合体蛋白磷酸化与去磷酸化有关。
用非洲爪蟾的成熟卵母细胞研究核膜的优势?
非洲爪蟾的成熟卵母细胞处于第二次减数分裂中期。此时的卵母细胞体积很大, 直径可达1mm, 其中储存了大量的营养物质, 为受精后快速分裂做准备。一个成熟的卵母细胞所储备的原料可供形成1000-10000个细胞核。
核孔复合体
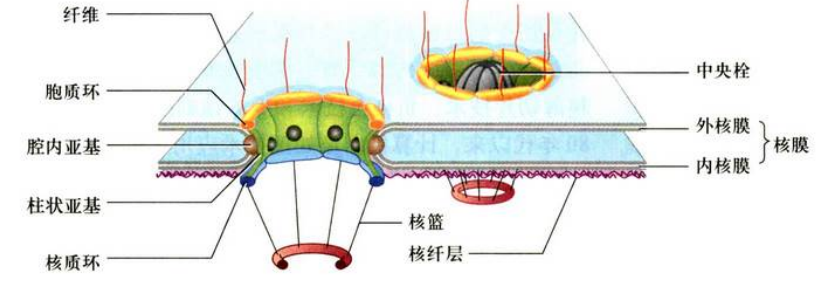
核孔复合体(nuclear pore complex, NPC): 在核孔上镶嵌着一种复杂的结构, 叫做核孔复合体, 主要由蛋白质构成, 是间期细胞核表面普遍存在的结构。核孔复合体在核被膜上的数量、分布密度与分布形式随细胞类型、细胞核的功能状态的不同而有很大的差异。转录功能越活跃的细胞, 核孔复合体的数量越多。
结构模型
研究核孔复合体形态结构的3种方法: 树脂包埋超薄切片技术;负染色技术;冷冻蚀刻技术。核孔复合体的结构模型:
核孔复合体由四部分组成: 胞质环、核质环、中央栓、辐
- 胞质环(cytoplasmic ring): 胞质环位于核孔边缘细胞质基质一侧, 又称外环。环上有八条纤维对称分布伸向细胞质。
- 核质环(nuclear ring): 核质环位于核孔边缘细胞核基质一侧, 又称内环。环上有八条纤维对称分布伸向细胞核基质, 其末端形成由八个蛋白质构成的环, 就像一个捕鱼网, 称为核篮。
- 中央栓(central plug): 中央栓位于核孔中心, 呈颗粒状或者棒状, 在核质交换过程中起重要作用。
- 辐(spoke): 辐由核孔边缘伸向核孔中心, 呈辐射状八重对称, 由三个结构域构成, 包括柱状亚单位、腔内亚单位和环带亚单位。
- 柱状亚单位: 主要的区域位于核孔边缘, 连接内、外环, 起支撑作用;
- 腔内亚单位: 在这个结构域之外, 接触核膜的部分;
- 环带亚单位: 在柱状亚单位之内, 靠近核孔复合体中心的部分, 由8个颗粒状或棒状, 所以又称中心颗粒。
核孔复合体在核膜的轴向呈辐射状八重对称;
核孔复合体在核膜的平行向不对称, 即胞质面与核质面不对称, 这与其功能的不对称性相一致。
组成成分
核孔复合体主要由蛋白质构成, 其中gp210与p62是最具有代表性的两种类型。目前倾向于把所有的核孔复合体蛋白同一成为核孔蛋白(Nup)。
- gp210: 一类结构性跨膜蛋白, 是第一个被鉴定出来的核孔复合体蛋白, 糖基化修饰位点在天冬氨酸上, 所以是N-连接的甘露糖残基, 与coA有较强的结合作用。
- 功能: 介导核孔复合体与核被膜的连接, 将核孔复合体锚定在”孔膜区”, 从而为孔复合体组装提供一个起始位点;在内、外核膜融合形成核孔中起重要作用;在核孔复合体的核质交换功能活动中起一定作用, 如gp210核周间隙内肽段的抗体能够降低蛋白质入核转运的速度。
- p62: 功能性的核孔复合体蛋白。是O-连接N-乙酰葡萄糖胺残基寡糖修饰。主要分为两个结构域: 疏水性N端区: 具有FXFG(F: 苯丙氨酸, X: 任意氨基酸, G: 甘氨酸), 可能直接参与物质交换;C端区: 具有疏水性的7肽重复序列, 类似于一些纤维蛋白的杆状区, 适合形成α螺旋, 这个区域可能通过卷曲螺旋与其他核孔复合体相互作用, 从而将p62分子稳定到核孔复合体上, 为其N端进行核质交换活动提供支持。
功能
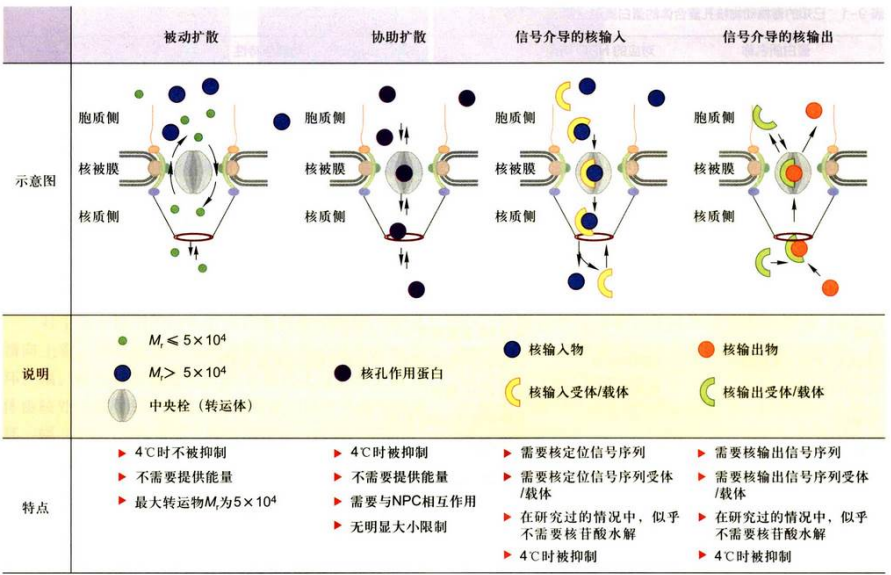
核孔复合体是一种特殊的跨膜运输蛋白复合体, 并且是一个双功能、双向性的亲水性核质交换通道。双功能表现在它有两种运输方式:被动扩散与主动运输。双向性表现在既介导蛋白质的入核转运, 又介导RNA、
- 通过核孔复合体的被动扩散: 核孔复合体作为被动扩散的亲水通道, 其有效直径为9~10nm, 有的可达12.5nm,即离子、小分子以及直径在10nm以下的物质原则上可以自由通过。
- 核孔复合体的主动运输: 主要是通过核孔复合体的主动运输完成的, 具有高度的选择性, 并且是双向的。
核孔复合体主动运输的选择性表现:
- 对运输颗粒大小的限制。主动运输的功能直径比被动运输大, 为10~20 nm, 甚至可达26nm。像核糖体亚基那样大的RNP也可以通过核孔复合体从核内运输到细胞质中, 表明核孔复合体的有效直径的大小是可调节的。
- 通过核孔复合体的主动运输是一个, 信号识别与载体介导的过程, 需要消耗ATP能量, 并表现出饱和动力学特征。
- 通过核孔复合体的主动运输具有双向性, 即核输入与核输出, 它既能把复制、转录、染色体构建和核糖体亚单位组装等所需要的各种因子运输到核内;同时又能将翻译所需的RNA、组装好的核糖体亚单位从核内运送到细胞质。
亲核蛋白(karyophilic protein): 是指在细胞质内合成后, 需要或能够进入细胞核内发挥功能的一类蛋白质。亲核蛋白一般都含有特殊的氮基酸序列, 这些内含的特殊短肽保证了整个蛋白质能够通过核孔复合体被转运到细胞核内。
核定位信号(nuiclear localization signal,NLS): 一段含有特殊的氮基酸序列片段, 富含碱性氨基酸残基, 作用是帮助和蛋白进入细胞核。这种入核信号特点: ①由含水的核孔通道来鉴别;②核定位信号是蛋白质的永久性部分, 在进入细胞核后, 并不被切除, 可以反复使用, 有利于细胞分裂后核蛋白重新输入核。
亲核蛋白的入核转运步骤:
- 亲核蛋白通过NLS识别importinα, 与可溶性NLS受体importinα/ improtinβ异二聚体结合, 形成转运复合物。
- 在importinβ的介导下, 转运复合物与核孔复合体的胞质纤维结合。
- 转运复合物通过改变构象的核孔复合体从胞质面被转移到核质面。
- 转运复合物在核质面与Ran-GTP结合, 并导致复合物解离, 亲核蛋白释放。
- 受体的亚基与结合的Ran返回胞质, 在胞质内Ran-GTP水解形成Ran-GDP并与importinβ解离, Ran-GDP返回核内再转换成Ran-GTP状态。
亲核蛋白单向输入的原因:
- 核孔复合体本身结构不对称;
- 胞质与核质中的 Ran-GTP 的浓度梯度;
- 亲核蛋白入核与胞质中蛋白质分选的差异性:
亲核蛋白由核孔复合体介导入核,受到核孔复合体调控,而非载体蛋白;亲核蛋白的信号序列NLS在入核后不被切除;亲核蛋白以完全天然的构象入核,无需分子伴侣协助其解折叠。
RNA and RNPs核输出:
- 由RNA pol I转录形成的rRNA, 需要在核内装配形成核糖核蛋白亚单位(RNPs), 再经由核孔复合体转运至细胞质, 此过程消耗代谢能量。
- 由RNA pol II转录形成的hnRNA, 需要经过转录后加工, 即3’端加尾, 5’端加帽以及剪接等过程后由核孔复合体输出。
- 由RNA pol III转录形成的5S rRNA、tRNA, 经一种蛋白介导转运输出。
核纤层
核纤层(nuclear lamina): 存在于间期细胞核中, 紧贴内层核膜下, 是一层由纤维蛋白构成的网络结构, 分布于内层核膜和染色质之间并与染色质及核骨架相连, 对核被膜起支撑作用。在细胞分裂过程中对核被膜的破裂和重建起调节作用。
哺乳动物体细胞的核纤层主要有3种: laminA、laminB和laminC。核纤层蛋白A和C是同一个基因的不同拼接体, 前面566个氨基酸完全相同, 但是C端序列不一样。核纤层蛋白A和C的表达具有组织与发育时期的特异性, 但核纤层蛋白B则在哺乳动物的所有细胞中都表达。
核纤层的功能:
- 结构支撑功能: 核纤层蛋白形成骨架结构支撑于核被膜的内侧, 使得核被膜能起到细胞核与细胞质之间的隔离与信息交换功能。核纤层的骨架功能还得以使细胞核维持正常的形状与大小。
- 调节基因表达: 果蝇细胞中基因组范围的研究结果表明, 沉默基因更倾向于分布于核纤层附近, 异染色质更易与核纤层结合, 而且核纤层附近的染色质的乙酰化水平较低。然而, 在酵母细胞中活跃转录的基因也分布于核纤层附近, 它们常与核孔复合体结合。所以, 核纤层与基因表达的确切关系还不是非常清楚。很可能在不同物种的细胞中, 甚至不同组织的细胞中情况不一样。
- 调节DNA修复: 研究表明, laminA核纤层蛋白是双链DNA断裂修复必需的。核纤层蛋白功能异常病人细胞中的基因组变得不稳定, DNA修复反应滞后, 端粒变短。
- 与细胞周期的关系: 细胞分裂过程中, 核纤层蛋白解聚成可溶的单体或与崩解后的核被膜相结合。新核形成时, 核被膜与染色质结合的同时, 核纤层也最后重新形成。
核纤层蛋白基因突变导致的疾病:
Hutchinson-Gilford早老综合征是由于LaminA的C端的50个氨基酸的缺失;
非典型性Werner综合征是由于A57P、R133L和L140R的氨基酸改变;核纤层蛋白B受体功能改变引起Pelger-huet异常疾病。
核纤层在细胞分裂过程中的变化:
分裂前期, 核膜崩解, 核纤层解聚, A型核纤层蛋白以可溶性单体形式弥散于胞质中, B型核纤层蛋白则与核膜小泡保持结合状态;
分裂末期, 核膜重现, 核纤层在染色体周围重新装配;
核纤层蛋白的磷酸化与去磷酸化可能是核纤层结构动态变化的调节因素。
染色质
染色质(chromatin): 是细胞核中能被碱性染料强烈着色的物质, 故名染色质。染色质由DNA、组蛋白、非组蛋白及少量RNA组成, 是间期细胞遗传物质存在的形式, 是遗传物质的载体。
染色体(chromosome): 是指细胞在有丝分裂或减数分裂过程中, 由染色质聚缩而成的棒状结构。在化学本质上和染色质没有差别, 只是构型上不同, 是遗传物质在细胞周期不同阶段的不同表现形式。不同生物的细胞中含有不同数目的染色体。
染色体与染色质的异同之处: 染色体和染色质在化学本质上没有差异, 只是在构型上不同,是遗传物质在细胞周期不同阶段的不同表现形式。在真核细胞的细胞周期中, 大部分时间是以染色质的形态而存在的。
染色质DNA
基因组大小比较
基因组(genome): 某一生物的细胞中储存于单倍染色体组中的总遗传物质。
基因组DNA类型
生物基因组DNA分类:
- 蛋白编码序列;
- 编码rRNA、tRNA、snRNA和组蛋白的串联重复序列;
- 含有重复序列的DNA;
- 未分类的间隔DNA。
高度重复DNA序列分类:
- 卫星DNA(satellite DNA): 重复单位长5~100 bp, 不同物种重复单位碱基组成不同, 一个物种也可能含有不同的卫星DNA序列。主要分布在染色体着丝粒部位。如人类染色体着丝粒区的α-卫星DNA家族, 其功能不明。
- 小卫星DNA(minisatellite DNA): 重复单位长12-100bp, 重复3 000次之多。又称数量可变的串联重复序列。每个小卫星区重复序列的拷贝数是高度可变的, 常用于DNA指纹技术做个体鉴定。研究发现小卫星序列的改变可以影响邻近基因的表达, 基因的异常表达会导致一系列不良后效应。
- 微卫星DNA(mnicrosatellite DNA): 重复单位序列最短, 只有1~5bp, 串联成簇长度50-100bp。人类基因组中至少有30 000个不同的微卫星位点, 具高度多态性, 在不同个体间有明显差别, 但在遗传上却是高度保守的。可作为重要的遗传标记, 用于构建遗传图谱及个体鉴定等。
染色质蛋白
组蛋白
组蛋白(histone): 组蛋白是真核生物染色体的基本结构蛋白, 是一类进化上保守小分子碱性蛋白质,与DNA结合没有序列特异性。有五种类型: H1、H2A、H2B、H3、H4, 它们富含带正电荷的碱性氨基酸,能够同DNA中带负电荷的磷酸基团相互作用。5种组蛋白在功能上分为两组: ①核小体组蛋白(H3、H4、H2A、H2B);②H1组蛋白。
非组蛋白
非组蛋白(nonhistone): 是指细胞核中组蛋白以外的酸性蛋白质, 与特定的DNA序列或组蛋白相结合。非组蛋白不仅包括以DNA作为底物的酶, 还包括作用于组蛋白的一些酶, 此外还有DNA结合蛋白、组蛋白结合蛋白和调节蛋白。
利用非组蛋白与特异DNA序列亲和的特点, 通过凝胶延滞实验(gel retardation assay)可以在细胞抽提物中进行检测。首先制备一段带有放射性标记的已知特异序列的DNA,将要检测的细胞抽提物与标记DNA混合, 进行凝胶电泳。未结合蛋白质的自由DNA在凝胶上迁移最快, 而与蛋白质结合的DNA迁移慢, 一般结合的蛋白质分子越大, DNA分子的延滞现象越明显, 然后通过放射性自显影分析, 即可发现一系列DNA带谱,每条带分别代表不同的DNA-蛋白质复合物。每条带相对应的结合蛋白随后再通过细胞抽提物组分分离方法被进一步分开。
非组蛋白具有以下特性:
- 非组蛋白具有多样性:非组蛋白占染色质蛋白的 60%~70%,不同组织细胞中其种类和数量都不相同,代谢周转快。
- 识别 DNA 具有特异性:能识别特异的 DNA 序列,识别信息来源于DNA 核苷酸序列本身,识别位点存在于 DNA 双螺旋的大沟部分,识别与结合靠氢键和离子健。
- 具有功能多样性:具有多方面的重要功能,包括基因表达的调控和染色质高级结构的形成。
非组蛋白的结构模式:
模式一: α螺旋-转角-α螺旋(helix-turn-helix motif)最简单最普遍;两个α螺旋组成一个β转角;羧基端的螺旋负责识别DNA序列
模式二: 锌指模式;
模式三: 亮氨酸拉链模式;
模式四: 螺旋-环-螺旋模式;
模式五: HMG框结构模式
DNA与蛋白质结合的验证:
- 凝胶延滞实验: 是一种分离试验让大小不同的粒子在脉冲的作用下由凝胶的一端运动到另一端由与凝胶对不同粒子阻碍作用的大小受粒子本身大小所影响所以不同大小的粒子到达另一端的先后顺序不同从而达到分离效果。
- 染色质免疫沉淀实验(ChIP assay)
核小体
核小体(nucleosome): 是由DNA和组蛋白形成的染色质基本结构单位。每个核小体由146bp的DNA缠绕组蛋白八聚体形成。核小体核心颗粒之间通过50bp左右的连接DNA相连。H1结合在盘绕在八聚体上的DNA双链开口处。
核小体的发现
- 用温和的方法裂解细胞核得到核小体彼此连接的串珠状结构, 串珠的直径为11nm;
- 用非特异性微球菌核酸酶消化染色质时, 经过蔗糖梯度离心及琼脂糖凝胶电泳分析, 得到大量200bp或200bp整数倍的DNA片段;
- 应用X射线衍射、中子散射和电镜三维重建技术, 研究染色质结晶颗粒, 观察到八聚体现象;
- SV40微小染色体分析。
核小体的结构
- 每个核小体包括200bp左右的DNA和一个组蛋白八聚体以及一分子的组蛋白H1。
- 八聚体包括两分子的H2A·H2B和两分子的H3·H4。
- 147bp的DNA盘绕组蛋白八聚体1.75圈, H1结合额外20bp DNA, 锁住核小体DNA的进出端, 稳定核小体。
- 两个相邻核小体之间以连接DNA相连, 典型长度为60bp, 不同物种变化值为0~80bp不等。组蛋白密码(histone code): 组蛋白在翻译后的修饰过程中发生改变, 提供一种识别的标志, 为其他蛋白与DNA的结合产生协同或拮抗效应, 是一种动态转录调控成分, 称为组蛋白密码, 其中包括被修饰的氨基酸种类, 位置, 和修饰方式。
染色质组装
染色体高级结构
1.染色体高级结构的基础
- 组蛋白H1与核小体的结合对染色体高级结构的形成至关重要。H1的结合使DNA与核心组蛋白的缠绕更为紧密, 以介导形成染色体的高级结构, 并使其可接触性降低, 结构稳定。
- 核心组蛋白N端尾巴与相邻核小体间的相互作用对形成染色体高级结构至关重要。
2.染色体的二级结构
染色体的二级结构是指30nm螺线管结构, 以6个核小体为一圈, 旋转排列形成外径约30nm的螺线管。由10nm核小体串珠到30nm螺线管, DNA压缩了6倍。
3.染色体的三级结构
由螺旋管进一步螺旋化形成直径为0.4um的圆筒状结构, 称为超螺旋, 这是染色质的三级结构。
4.染色体的高级结构模型: 共压缩8400倍
- 多级螺旋模型: 30nm螺线管进一步盘区缠绕, 形成外径约400nm的超螺线管结构, 再由超螺线管组装形成染色单体。
- 绊环模型: 30nm螺线管进一步盘曲折叠, 形成DNA复制环, 每18个复制环平面放射状排列, 与核基质结合形成微带, 约106个微带沿纵轴排列形成染色单体。
染色质组装的前期过程
- H3·H4四聚体结合, 与新合成的裸露的DNA结合;
- 两个H2A·H2B异二聚体加入形成一个核心颗粒, H4上Lys5和Lys12被乙酰化;
- ATP创建一个规则的间距, 组蛋白去乙酰化, H1结合DNA伴随核小体折叠;
- 6个核小体组成一个螺旋或由其他的组装方式形成一个直径25~30nm的螺线管结构。
染色质组装的后期过程
- 染色质组装的多级螺旋模型
1
2
3
4
5flowchart LR
DNA-->核小体
核小体-->螺线管
螺线管-->超螺线管
超螺线管-->染色单体
- 染色质组装的放射环结构模型
1
2
3
4
5
6flowchart LR
DNA-->核小体
核小体-->螺线管
螺线管-->DNA复制环
DNA复制环-->微带
微带-->染色单体
放射环结构模型(scaffold radial loop structure model): 30 nm的染色线折叠成环, 沿染色体纵轴, 由中央向四周伸出, 构成放射环, 即染色体的骨架-放射环结构模型。
染色质类型
常染色质与异染色质
常染色质(euchromatin): 是指在分裂间期细胞内折叠、压缩程度较低, 形态较伸展, 用碱性染料染色时着色较浅的染色质。构成常染色质的DNA主要是单一序列DNA和中度重复序列DNA。常染色质并非所有基因都有转录活性。
异染色质(heterochromatin): 异染色质是指在分离间期细胞内折叠、压缩程度较高, 形态较收缩, 用碱性染料染色时着色较深的染色质。分为结构异染色质(组成型异染色质)和兼性异染色质。
结构异染色质(constitutive heterochromatin): 在整个细胞周期内都处于凝集状态的染色质, 即永久性的呈现异固缩的染色质被称为结构性异染色质。结构性异染色质含有高度重复的随体DNA, 分布于大多数染色体的着丝粒区、端粒和次缢痕处, 呈现C带染色。结构性异染色质的DNA主要是高度重复顺序,含有的基因相当少。
结构异染色质特征:
- 在中期染色体上多定位于着丝粒区、端粒、次缢痕及染色体臂的某些节段。
- 由相对简单、高度重复的DNA序列构成, 如卫星DNA。
- 具有显著的遗传惰性, 不转录也不编码蛋白质。
- 在复制行为上与染色质相比表现为晚复制、早聚缩。
- 占有较大部分核DNA, 在功能上参与染色质高级结构的形成, 导致染色质区间性, 作为核DNA的转座元件, 引起遗传变异。
兼性异染色质(facultative heterochromatin): 在某些细胞类型或一定的发育阶段, 原来的常染色质聚缩, 并丧失基因转录活性, 变为异染色质, 如巴氏小体。这类兼性异染色质的总量随不同细胞类型而变化, 一般胚胎细胞含量很少, 而高度特化的细胞含量较多。
染色体领域(chromosome territory): 间期核内的染色体并不是散乱分布的, 而是各自占据了一块特定的核区域称为染色体领域。这些区域的排列不定位是经过严格组织的, 并具有一定的动力学特征, 与基因的表达调控密切相关。
常染色质与异染色质之间的转变
- 基本原理: 异染色质或常染色质之间随着发育时期或细胞周期的变化而相互转化, 这种转变伴随着一些组蛋白与DNA修饰。
- 常见修饰
- 乙酰化: 通常促进异染色质转化为常染色质。
- 甲基化: 通常促进常染色质转化为异染色质。
活性染色质与非活性染色质
活性染色质(active chromatin): 是指具有转录活性的染色质。是由于核小体构象发生构型改变, 转变成疏松的染色质结构, 从而便于转录调控因子与顺式作用元件结合和RNA聚合酶在转录模板上滑动。
活性染色质的标志: H3 N端第4个赖氨酸的甲基化, 第9和14个赖氨酸的乙酰化以及第10个丝氨酸的磷酸化。
活性染色质分离方法: 有机汞亲和层析和二硫苏糖醇(DTT)洗脱: 当染色质结构处于疏松状态时, 利用核心组蛋白H3暴露出来的游离巯基与有机汞的亲和性。
非活性染色质(inactive chromatin):是指没有转录活性的染色质。非活性染色质的标志: H3N端第9个赖氨酸甲基化而不是乙酰化。
活性染色质对DNaseI 超敏感
DNaseⅠ超敏感位点(DNase I hypersensitive site): 是指用很低浓度的DNaseⅠ处理染色质, 切割首先发生的少数特异性位点, 这些特异性位点叫做DNaseⅠ超敏感位点。它是一段长100-200bp的DNA序列特异暴露的染色质区域。超敏感位点的存在是活性染色质的特点, 每个活跃表达的基因都有一个或几个超敏感位点, 大部分位于基因5’端启动子区域, 少部分位于其他部位如转录单位的下游。
核酸酶超敏感位点(nuclease-supersensitive site): 在染色质DNA中对DNaseⅠ表现出高度敏感的区域, 缺少核小体。这种区域通常很短, 最长也不过几百bp。该位点含有特异的DNA序列, 为特异性DNA结合蛋白所识别, 因此, 它参与了基因表达的调控。另外, 处于活性转录的区域对核酸酶也十分敏感。
活性染色质的蛋白组成特点:
- 很少有组蛋白H1与活性染色质结合;
- 活性染色质组蛋白乙酰化程度高;
- 核小体组蛋白H2B很少被磷酸化;
- 核小体组蛋白H2A高度保守;
- 组蛋白H3的变种H3.3只在活跃转录的染色质中出现;
- HMG蛋白是染色体非组蛋白中一组较丰富、不均一、富含电荷、较保守的蛋白质, HMG14和HMG17只存在于活性染色质中, 与DNA结合。
染色质的复制与表达
染色质的复制与修复
染色质的复制
- 两个过程
- DNA的复制;
- DNA与蛋白质组装形成染色质。
- 两种染色质组装途径
- 亲代转移: 在复制叉的移动期间, 父代的核小体核心颗粒与DNA分离, 在该段DNA复制完成时, 父代的核小体核心颗粒直接转移到两条子链DNA的一条上。
- 重新合成: 染色质组装因子利用刚合成的、乙酰化的组蛋白介导核小体在复制DNA上组装。
染色质的修复与基因组稳定性
- 高度浓缩的染色质结构对基因组DNA(染色质DNA)具有重要的保护作用;
- 细胞中存在一整套的DNA修复机器以应对各种各样的DNA突变, 使基因组稳定;
- 修复好的DNA必须及时地与组蛋白结合, 组装成染色质。
染色质的激活与失活
染色质结构改变与基因活化的关系研究
- 如何形成活性染色质, 以便RNA聚合酶起始转录;
- 具有转录活性的染色质结构域如何与周围的非活性区域隔离;
- RNA聚合酶如何通过与染色质模板结合进行转录。
染色质的激活
1.DNA结构与核小体相位的变化
- DNA构象变化: 第一、调控蛋白结合到染色质特定位点时, 易引发染色质二级结构的改变, 使其他结合位点更易或更难与调节蛋白结合。第二、拓扑异构酶能调整DNA双螺旋变化, 使之超螺旋化或者松弛。
- 核小体定位于DNA特定位点: 第一、基因关键调控元件(增强子和启动子)位于核小体颗粒之外, 使之便于与转录因子结合。第二、基因调控元件位于盘绕核心组蛋白的DNA上, 则增强子和启动子两种调控元件通过转录因子被联系起来。
2.组蛋白的修饰
- 基本修饰: 第一、核心组蛋白的赖氨酸残基乙酰化, 激活染色质。第二、组蛋白H1的磷酸化, 导致染色质疏松。
- 修饰意义: 第一、改变染色质的结构, 直接影响转录活性。第二、核小体表面发生改变, 使调控蛋白容易和染色质结合, 间接影响转录活性。
3.HMG蛋白的影响
具有HMG结构域的蛋白可特异性识别DNA, 与DNA折叠和DNA-蛋白质复合物高级结构的形成有关。
染色质的失活
1.X染色体失活
雌性哺乳动物中失活的X染色体上的H4组蛋白不发生乙酰化, 而且DNA高度甲基化, 不表达。
2.位置花斑效应
位置花斑效应(position effect variegation): 位置花斑效应是指活性基因转位到异染色质区附近时会失活的现象。
隔离子(insulator): 是指位于抑制状态与活化状态的染色质结构域之间、能防止不同状态的染色质结构域的结构特征向两侧扩展的染色质DNA序列。作用是作为染色质定向形成的起始位点, 提供拓扑隔离区。隔离子的作用:
- 作为异染色质定向形成的起始位点;
- 作为结构域两端的锚定点, 提供拓扑隔离区, 使结构域外的增强子成分不能进入;
- 涉及追踪机制, 远端增强子处组成的复合体沿染色质模板运动直到启动子, 而隔离子可阻止这个复合体超越正常作用范围。
染色质与基因表达调控
以染色质为模板的转录
- 真核细胞中基因转录的模板是染色质而不是裸露的DNA, 染色质是否处于活化状态是决定RNA聚合酶能否有效行使转录功能的关键。
- 转录起始伴随着染色质上某一基因调节序列内部或者周围的结构改变。
转录因子介导的基因表达调控
转录因子(transcription factor): 是指能特异性识别DNA序列并与之结合从而对转录进行调节的蛋白质。转录因子从功能上可分为两类:通用转录因子和特异转录因子。
转录因子分类:
- 通用蛋白因子(general transcription factor): 与结合RNA聚合酶的核心启动子位点结合。
- 特异性蛋白因子(specific transcription factor): 与特异基因的各种调控位点结合, 促进或阻遏基因的转录。
转录因子的基本结构域:
- DNA结合域。DNA结合基序: 锌指、螺旋-转角-螺旋(HTH)、亮氨酸拉链和螺旋-环-螺旋(HLH)。
- 激活或抑制结构域。
DNA甲基化介导的基因表达调控
DNA甲基化的影响:
- DNA甲基化与基因表达的阻遏有关, 是使DNA转入持久遏制状态的重要条件;
- DNA甲基化在基因组印记中具有重要作用。
甲基化抑制转录的两种方式:
- 干扰转录因子对DNA结合位点的识别;
- 将转录激活因子识别的DNA序列转换为转录抑制因子的结合位点。
组蛋白修饰介导的基因表达调控
组蛋白的乙酰化、甲基化以及磷酸化等修饰改变了染色质的结构, 调控基因的表达。
染色质与表观遗传
表观遗传与遗传
遗传学与表观遗传的区别:
- 遗传学的变化是通过DNA序列的突变实现的, 通过生殖细胞得以遗传。
- 表观遗传的变化是通过组蛋白和DNA的不同修饰而实现的, 只在体细胞中出现。
表观遗传
表观遗传(epigenetics): 表观遗传是指在DNA序列不发生变化的情况下, 在环境条件的影响下, DNA或组蛋白发生不同修饰而使基因表达发生的可遗传性变化。
改变染色体结构的主要途径:
- 各种组蛋白修饰共同构成组蛋白密码子;
- 染色质重塑;
- 组蛋白异型体;
- DNA甲基化;
- 非编码RNA的作用。
表观遗传的本质是由DNA修饰、多种组蛋白修饰、染色质重塑因子及组蛋白伴侣等因素而引起的遗传学变化。
染色体
染色体的形态结构
中期染色体分类
- 中着丝粒染色体(metacentric chromosome): 着丝粒位于染色体的中部, 两臂长度相等或大致相等;
- 亚中着丝粒染色体(submetacentric chromosome): 着丝粒偏离中部, 染色体的两个臂长短不一;
- 近端着丝粒染色体(acrocentric chromosome): 着丝粒靠近染色体的一端, 长臂极长, 短臂极短;
- 端着丝粒染色体(telocentric chromosome): 着丝粒位于染色体的末端, 只有一个长臂。
着丝粒与动粒
着丝粒(centromere):是染色体中连接两个染色单体, 并将染色单体分为两臂: 短臂和长臂的结构。由于着丝粒的染色质较细、内缢, 又叫主缢痕。此处 DNA 具高度重复, 为碱性染料所深染。着丝粒有两个基本的功能:在有丝分裂前将两条姐妹染色单体结合在一起,第二个功能为动粒装配提供结合位点。一个染色体只有一个着丝粒。
动粒(kinetochore):是由着丝粒结合蛋白在有丝分裂期间特别装配起来的、附着于主缢痕外侧的圆盘状结构,内层与着丝粒结合,外层与动力微管结合。每一个中期染色体含有两个动粒,位于着丝粒的两侧。动粒与纺锤体微管相连,确保染色体能够平均分配到子细胞中。
着丝粒的基本组成:
- 动粒结构域: 包括与着丝粒中央结构域相联系的内板、中间间隙、外板和纤维冠。
- 中央结构域: 由串联重复的卫星DNA组成, 是着丝粒区的主体。
- 配对结构域: 代表中期姐妹染色单体相互作用的位点。
次缢痕
次缢痕(secondary constriction): 是指除主缢痕外, 染色体上其他的浅染缢缩部位, 它的数目、位置和大小是某些染色体所特有的形态特征, 因此也可以作为鉴定染可以作为鉴定染色体的标记。
核仁组织区(NOR)
核仁组织区(nucleolar organizing region, NOR): NOR位于染色体的次缢痕部位, 但并非所有次缢痕都是NOR。染色体NOR是rRNA基因所在部位(5S rRNA基因除外), 与间期细胞核仁形成有关。
随体
随体(satellite): 是指位于染色体末端的球形染色体节段, 通过次缢痕区与染色体主体部分相连。它是识别染色体的重要形态特征之一, 有随体的染色体称为sat染色体。根据随提在染色体上的位置, 可分为两大类: 端随体和中间随体。
端粒
端粒(telomere): 是指染色体两端由富含G的短的串联重复序列DNA组成的特化结构, 多以单链形式存在, 伸展到染色体的3’端。人类端粒序列为5’-TTAGGG-3’。一个基因组内的所有端粒都是由相同的重复序列组成, 有物种特异性。端粒能够保护染色体末端, 避免末端被降解或重组, 维持染色体的完整性和独立性, 端粒的长度与细胞及生物个体的寿命有关。
染色体的功能元件
确保染色体复制和稳定遗传应具备的3个元件:
- 至少一个DNA复制起点, 确保染色体在细胞周期中能够自我复制, 维持染色体在细胞世代传递中的连续性;
- 一个着丝粒, 使细胞分裂时已完成复制的染色体能平均分配到子细胞中;
- 最后, 在染色体的两个末端必须有端粒, 保持染色体的独立性和稳定性。
“人造微小染色体”(artificial minichromosome): 采用分子克隆技术把真核细胞染色体的复制起点、着丝粒和端粒这3种DNA关键序列分别克隆成功, 并把它们互相搭配或改造而构成人造微小染色体, 可以像天然染色体一样在寄主细胞中稳定复制和遗传。
复制起点序列
自主复制序列(autonomous replicating sequence, ARS序列): 又叫复制起点序列(replicationorigin sequence), 这种序列是染色体正常起始复制所必需的顺式作用元件。所有的ARS的DNA均有一段保守序列, 上下游各200个bp左右的序列是维持ARS功能所必需的。原核生物只有一个复制起点。真核生物有多个复制起点。
作用:复制起点序列是细胞周期中 DNA 复制的起点,维系染色体在细胞分裂过程中的连续性。
着丝粒序列
着丝粒(centromere): 是染色体中连接两个染色单体, 并将染色单体分为两臂: 短臂和长臂的结构。由于着丝粒的染色质较细、内缢, 又叫主缢痕。此处DNA具高度重复, 为碱性染料所深染。着丝粒有两个基本的功能: 在有丝分裂前将两条姐妹染色单体结合在一起, 第二个功能为动粒装配提供结合位点。一个染色体只有一个着丝粒。
端粒序列
端粒(telomere): 是指染色体两端由富含G的短的串联重复序列DNA组成的特化结构, 多以单链形式存在, 伸展到染色体的3’端。人类端粒序列为5’-TTAGGG-3’。一个基因组内的所有端粒都是由相同的重复序列组成, 有物种特异性。
作用:
- 端粒能够保护染色体末端,避免末端被降解或重组。
- 端粒的生物学作用在于维持染色体的完整性和独立性。
- 端粒的长度与细胞及生物个体的寿命有关。
- 可能还与染色体在核内的空间排布等有关。
染色体带型
核型(karyotype): 是指染色体组在有丝分裂中期的表型, 是染色体数目、大小、形态特征的总和。核型分析是指在对染色体进行测量计算的基础上, 进行分组、排队、配对并进行形态分析的过程。核型分析对于探讨人类遗传病的机制、物种亲缘关系、远缘杂种的鉴定等都有重要意义。核型模式图(idiogram): 将一个染色体组的全部染色体逐个按其特征绘制下来, 再按长短、形态等特征排列起来的图象。
核型分析研究方法:
- 低渗处理技术;
- 秋水仙素富集中期细胞分裂相;
- 植物凝集素(PHA)刺激血淋巴细胞转化、分裂。
染色体显带技术: 经物理、化学因素处理后, 再用染料对染色体进行分化染色, 使其呈现特定的深浅不同带纹(band)的方法。
- 一类是产生的染色带分布在整过染色体长度上如: G、Q和R带,
- 一类是局部性显带, 它只能使少数特定区域显带, 如C、T和N带
特殊染色体
巨大染色体(giant chromosome): 在某些生物的细胞中, 特别是在发育的某些阶段, 可以观察到一些特殊的体积很大的染色体, 它们的特点是体积巨大、细胞核和整个细胞体积也大。主要包括多线染色体和灯刷染色体。)
多线染色体
多线染色体(polytene chromosome): 是指有丝分裂过程中, DNA多次复制但是细胞不分裂而使多股子染色单体平行排列的染色体结构。①产生的子染色体并行排列, 且体细胞内同源染色体配对, 紧密结合在一起从而阻止染色质纤维进一步聚缩, 形成体积很大的多线染色体。②多线化的细胞处于永久间期, 并且体积也相应增大。
灯刷染色体
灯刷染色体(lampbrush chromosome): 是指卵母细胞进行减数分裂第一次分裂时停留在双线期, 同源染色体未解除分离而形成二价体的染色体结构, 形似灯刷。灯刷染色体的侧环是RNA活跃转录的区域, 灯刷染色体合成的RNA主要为前体mRNA,有些类型的mRNA可翻译成蛋白质, 有些mRNA与蛋白质结合, 暂不翻译而储存在卵母细胞中。
核仁与核体
核仁(nucleolus): 是真核细胞有丝分裂间期细胞核内最显著的小球型结构, 是细胞核中rRNA的合成、加工和核糖体亚单位的组装场所, 在细胞周期中有规律的消失与重现。蛋白质合成旺盛、活跃生长的细胞的核仁大。
核仁的结构
纤维中心(fibrillar center, FC): FC是包埋于颗粒组分内的一个或几个电子密度较低的结构区域, 染色较浅。内含rDNA与RNA聚合酶Ⅰ和转录的结合因子。FC中的染色质不能形成核小体, 也没有组蛋白的存在, 但存在嗜银蛋白。通常认为FC代表染色体NOR在间期核的副本。
致密纤维组分(dense fibrillar component, DFC): DFC是包埋于颗粒组分内的电子密度较高的结构区域, 呈环形或半月形包围FC, 由致密纤维构成, 通常看不到颗粒。rDNA在FC于DFC交界处转录为pre-rRNA。
颗粒组分(granular component, GC): 在代谢活跃的细胞的核仁中, 颗粒组分是核仁的主要结构。由直径15~20nm的RNP构成, 可被蛋白酶和RNase消化。这些颗粒是正在加工、成熟的核糖体亚单位前体颗粒。间期核中核仁的大小差异主要是由颗粒组分数量的差异造成的。
核仁基质(核仁骨架): 包括核仁周边染色质与核仁内染色质, 构成核仁组织区。
核仁相随染色质: 核仁中的无定形物质, FC、DFC、GC等均包埋其中。
转录主要发生在FC与DFC的交界处。
核仁的功能
- rRNA的合成、加工
- 核糖体的组装成熟
- mRNA的输出与降解
核仁的动态周期变化
有丝分裂前期
细胞进入有丝分裂, 核仁变形、变小, 随着染色质凝集, 核仁消失。
有丝分裂中期和后期
在中期和后期细胞中没有核仁。
有丝分裂末期
在有丝分裂末期,核仁重建。
核体
核体(nuclear body): 是指间期核内除染色质与核仁外, 在染色质之间的空间还含许多形态上不同的亚核结构域, 统称为核体。
核基质
核基质(nuclear matrix): 是指真核细胞的核内除染色质、核膜与核仁外, 以蛋白质成分为主的网架结构。参与和支持DNA的各种功能, 包括DNA复制、转录、加工、接受外部信号以及维持染色质的结构等, 作用方式主要是提供作用位点。
核骨架(nuclear skeleton): 狭义上是指核基质;广义上包括核基质、核纤层以及染色体骨架。核骨架是存在于真核细胞细胞核中的, 由蛋白质构成的空间网络结构。核骨架的主要成分为非组蛋白构成的纤维蛋白, 含有少量RNA。核骨架与DNA复制、基因表达及染色体的组装与构建有密切关系。核骨架的功能:
1.核骨架与DNA复制
DNA聚合酶结合与核骨架上并被激活, 核骨架是DNA复制的空间支架;DNA复制起始位点是MAR。
2.核骨架与基因表达
具有转录活性的基因都结合在核骨架上;
RNA聚合酶在核骨架上具有结合位点;
RNA的合成是在核骨架上进行的, 基因只有结合在核骨架上才能转录。
3.与染色体构建有关
一般认为核骨架和染色质骨架为同一类物质, 30nm的染色质纤维就是结合在核骨架上, 形成放射环的结构, 在分裂期进一步包装成光学显微镜下可见的染色体。
染色体支架(chromosomal scaffold): 染色体中由非组蛋白极成的结极支架。染色体支架与核骨架二者有相同的组分, 核骨架的某些结极组分可能在分裂期中转变为染色体支架。
考点补充
名词补充