细胞生物学 第十章 核糖体
1
2
3
4
5
6
7
8
9 ____ __
/\ _`\ __/\ \
\ \ \L\ \/\_\ \ \____ ___ ____ ___ ___ ___ __
\ \ , /\/\ \ \ '__`\ / __`\ /',__\ / __`\ /' __` __`\ /'__`\
\ \ \\ \\ \ \ \ \L\ \/\ \L\ \/\__, `\/\ \L\ \/\ \/\ \/\ \/\ __/
\ \_\ \_\ \_\ \_,__/\ \____/\/\____/\ \____/\ \_\ \_\ \_\ \____\
\/_/\/ /\/_/\/___/ \/___/ \/___/ \/___/ \/_/\/_/\/_/\/____/
核糖体的类型与结构
核糖体的基本类型与化学组成
两种基本类型
- 原核细胞核糖体
- 真核细胞核糖体
- 细胞质核糖体
- 线粒体核糖体
- 叶绿体核糖体
两种核糖体的比较
| 化学组成 | 原核细胞核糖体(70S) | 原核细胞核糖体(70S) | 真核细胞核糖体(80S) | 真核细胞核糖体(80S) |
|---|---|---|---|---|
| 化学组成 | 小亚基 | 大亚基 | 小亚基 | 大亚基 |
| 沉降系数 | 30S | 50S | 40S | 60S |
| 亚基蛋白数 | 21 | 34 | 33 | 49 |
| rRNA | 16S | 23S、5S | 18S | 25~28S、5.8S、5S |
核糖体的结构
- rRNA: 构成核糖体的核心, 决定核糖体形态, 催化蛋白肽键形成。
- 蛋白质: 定位于核糖体的表面或者填充于rRNA间的缝隙, 稳定rRNA。
- 核糖体结构特点:
- 每个核糖体含有4个RNA分子的结合位点, 其中1个位点供mRNA结合, 3个位点供tRNA分子结合;
- 在核糖体大小亚基结合面, 特别是mRNA和tRNA结合处, 无核糖体蛋白分布;
- 催化肽键形成的活性位点由RNA组成;
- 大多数核糖体蛋白有一个球形结构域和伸展的尾部, 球形结构域分布于核糖体表面, 多肽链尾部伸入核糖体内折叠的rRNA分子中。
核糖体蛋白质与rRNA的功能
核糖体上重要位点
- mRNA结合位点: 蛋白质的起始合成首先需要mRNA与小亚基结合。
- A位点: 与新掺入的氨酰-tRNA结合的位点——氨酰基位点。
- P位点: 与延伸中的肽酰-tRNA结合的位点——肽酰基位点。
- E位点: 脱氨酰tRNA离开A位点到完全释放的一个位点。
- 延伸因子结合位点: 与肽酰tRNA从A位点转移到P位点有关的转移酶(即延伸因子EF-G)的结合位点。
- 肽酰转移酶催化位点。
rRNA的主要功能
- 具有肽酰转移酶的活性;
- 为tRNA提供结合位点(A位点、P位点和E位点);
- 为多种蛋白质合成因子提供结合位点;
- 在蛋白质合成起始时参与同mRNA选择性地结合及在肽链的延伸中与mRNA结合。(三)r蛋白的功能的推测
- 对rRNA折叠成有功能的三维结构是十分重要的;
- 在蛋白质合成中, 某些r蛋白可能对核糖体的构象起”微调”作用。
多核糖体与蛋白质的合成
多核糖体
多核糖体(polyribosome或polysome): 在合成蛋白质时, 一条mRNA串联多个核糖体, 每个核糖体可合成一条多肽链, 这样的核糖体称为多聚核糖体。以这种形式进行多肽合成, 对mRNA的利用及对其数量的调控更为经济和有效, 在相同mRNA数条件下能大大提高多肽的合成效率。
原核细胞中, 在mRNA合成的同时, 核糖体就结合到mRNA上, 即DNA转录成mRNA和mRNA翻译成蛋白质这两个生命活动是同时并几乎在同一部位进行的, 所分离的多核糖体常常与DNA结合在一起。真核细胞中, 多核糖体或附着在内质网上, 或游离在细胞质基质中。
蛋白质的合成
主要是3个阶段: 肽链的起始、肽链的延伸和肽链的终止。
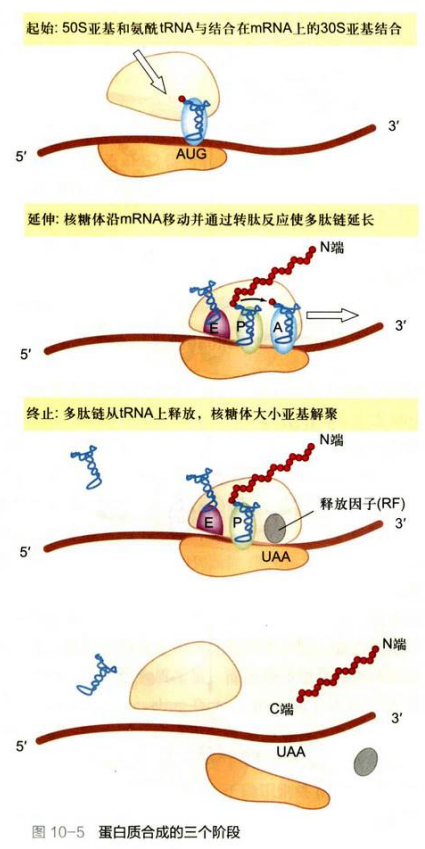
肽链的起始
蛋白质合成起始涉及mRNA、起始tRNA和核糖体小亚基间相互作用和装配。
- 30S小亚基与mRNA的结合
- 蛋白质合成起始阶段, mRNA通过AUG上游的SD序列与小亚基结合, 并使小亚基能精准的识别起始密码子AUG。
- 30S小亚基与mRNA的结合需要仅位于小亚基上的起始因子的帮助。在50S大亚基与30S小亚基结合后, 起始因子从小亚基上释放。
- IF1与30S亚基A位点结合, 协助30S亚基与mRNA的结合, 并防止氨酰-tRNA错误进入核糖体的A位点;IF2是一种GTP结合蛋白, 协助第一个氨酰-tRNA进入核糖体;
- IF3能防止核糖体50S大亚基提前与小亚基结合, 并有助于第一个氨酰-tRNA进入核糖体。
- 第一个氨酰-tRNA进入核糖体
- 完整起始复合物的装配
肽链的延伸
主要是4个步骤: 氨酰-tRNA进入核糖体A位点的选择、肽键的形成、转位和脱氨基-tRNA的释放。
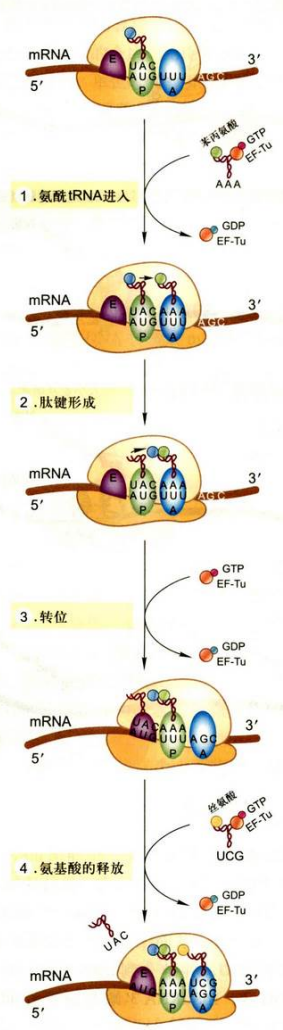
- 氨酰-tRNA进入核糖体A位点的选择
第二个氨酰-tRNA必须与有GTP的延伸因子EF-Tu结合形成复合物氨酰-tRNA·EF-Tu·GTP才能进入A位点。到位后, 结合在EF-Tu上的GTP水解, EF-Tu连同结合在一起的GDP离开核糖体, 被另一个因子EF-Ts介导生成EF-Tu·GTP。 - 肽键的形成
当核糖体的P位点与A位点都有tRNA时, 通过肽键的生成将两个氨基酸结合起来。具体来讲, 是A位点氨酰-tRNA氨基酸与P位点tRNA上氨基酸的羧基形成肽键。这一反应由肽酰转移酶催化, 该酶是核糖体大亚基rRNA, 活性位点位于23SrRNA结构域V的中央环。 - 转位
即核糖体沿着mRNA分子的5’→3’方向移动3个核苷酸。 - 脱氨酰-tRNA的释放
肽链的终止
终止密码子UAA、UAG、UGA。
释放因子RF1可识别UAG或UAA, RF2可识别UGA或UAA。
核糖体与RNA世界
核糖体的本质是核酶
核酶(ribozyme): 核酶化学本质是核糖核酸(RNA), 具有酶的催化功能。核酶的作用底物可以是不同的分子, 也可以是同一RNA分子中的某些部位。与蛋白质酶相比, 核酶的催化效率较低, 是一种较为原始的催化酶。
作用:
- 可催化RNA、DNA的水解和连接;
- 催化mRNA的剪接;
- 催化RNA聚合酶反应, RNA的磷酸化、氨酰基化等;
- 肽酰转移酶合成肽链。
(二)RNA世界与生命起源
抗生素干扰病原微生物的代谢过程的原理(补充):
- 抑制细菌细胞壁的合成(如青霉素);
- 影响细菌细胞膜的通透性(如多粘菌素);
- 抑制菌体蛋白质的合成(如氯霉素、四环素);
- 抑制细菌核酸合成(如灰黄霉素);
抗生素是主要的蛋白质合成抑制剂: (分子中也有原话)
- 链霉素主要抑制起始tRNA和非起始tRNA与核糖体的结合, 导致肽链合成的提前终止。此外, 链霉素也可引起遗传密码子的错读。
- 嘌呤霉素具有与tRNA分子末端类似的结构, 能够同氨基酸结合, 代替氨酰化的tRNA同核糖体的A位点结合, 并参入到生长的肽链中, 因而导致蛋白质合成的终止并释放出C端含有嘌呤素的不成熟的多肽。
考点补充
名词补充