分子生物学 第十四章 RNA剪接
1
2
3
4
5
6
7
8
9 ____ __ __ ______ ___
/\ _`\ /\ \/\ \/\ _ \ /\_ \ __ __
\ \ \L\ \ \ `\\ \ \ \L\ \ ____ _____\//\ \ /\_\ ___ /\_\ ___ __
\ \ , /\ \ , ` \ \ __ \ /',__\/\ '__`\\ \ \ \/\ \ /'___\/\ \ /' _ `\ /'_ `\
\ \ \\ \\ \ \`\ \ \ \/\ \ /\__, `\ \ \L\ \\_\ \_\ \ \/\ \__/\ \ \/\ \/\ \/\ \L\ \
\ \_\ \_\ \_\ \_\ \_\ \_\ \/\____/\ \ ,__//\____\\ \_\ \____\\ \_\ \_\ \_\ \____ \
\/_/\/ /\/_/\/_/\/_/\/_/ \/___/ \ \ \/ \/____/ \/_/\/____/ \/_/\/_/\/_/\/___L\ \
\ \_\ /\____/
\/_/ \_/__/
本章概要
外显子(exon): 许多真核基因是嵌合的, 由一段编码区组成, 它们被另外一段段非编码区隔开了, 编码区称为外显子。术语”外显子”是指在成熟RNA中保留的任何序列片段, 不管这些序列片段是否为编码序列。
非编码的外显子包括:mRNA的5’端和3’端的非翻译序列;剪接后稳定的非编码RNA全长序列如X染色体失活调节因子Xist;有功能的RNA片段, 如小分子RNA(microRNA)。
内含子(intron): 许多真核基因是嵌合的, 由一段编码区组成, 它们被另外一段段非编码区隔开了, 非编码区称为内含子。
RNA剪接(RNA splicing): 是指从DNA模板链转录出的最初转录产物中除去内含子, 并将外显子连接起来形成一个连续的RNA分子的过程。
可变剪接(alternative splicing): 指不同剪接方式(选择不同的剪接位点组合), 从一个mRNA前体产生不同的mRNA剪接异构体的过程。通过这种方式, 一个基因可以产生多个多肽产物, 称为亚型。
机制: 仍然是两步转酯反应, 通过选择不同的剪接位点组合, 产生多种mRNA。
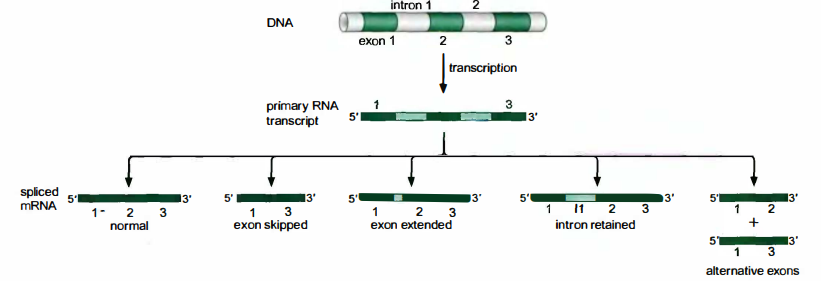
模式:
- 正常情况
- 外显子遗漏
- 外显子延伸
- 内含子保留
- 可变剪接外显子
生物学意义:
- 作为同一基因得到多种不同蛋白质的一种途径。
- 对某些基因起到开关作用。
RNA剪接的化学基础
RNA序列决定剪接位点
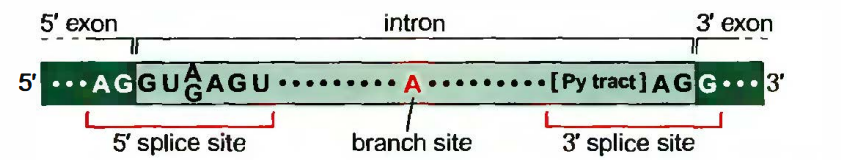
剪接的位点特征: 外显子和内含子边界, 即内含子的5’端是由称为5’剪接位点(5’splicesite)的序列标明的。在内含子3’端的内含子和外显子边界是由称为3剪接位点(3’splice site)的序列标明的。剪接所必需的第三个序列, 称为分支点(branchpoint site),或称分支点序列(branchpointsequence)。分支点全部位于内含子之内, 通常靠近3’端, 后面为多聚嘧啶区。
最保守的序列是内含子5’剪接位点的GU和3’剪接位点的AG及分支点的A。
当内含子两边的外显子连接时,内含子是以套索结构被除去的
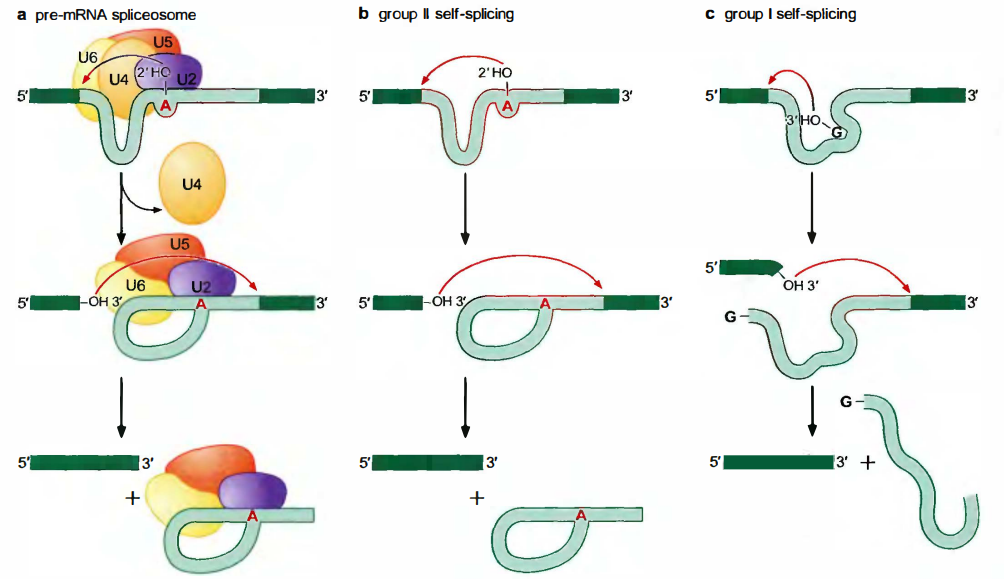
剪接是由两步连续的转酯反应(transesterification)完成的, 这样pre-mRNA中原来的一些磷酸二酯键断开, 在形成一些新的磷酸二酯键, 这个化学过程不需要消耗能量, 但正确组装和运行剪接机器需要ATP。剪接步骤:

第一步转酯反应是由分支点保守腺苷酸的2’羟基引发的。它作为亲核基团, 攻击5’剪接位点保守鸟苷酸的磷酰基团。结果, 外显子3’端的核糖与内含子5’端磷酸之间的磷酸二酯键断开, 而游离出来的5’磷酸与分支点保守腺苷酸的2’羟基连接。
在第二步转酯反应中, 5’外显子新游离出来的3’羟基反过来作为亲核基团, 攻击3’剪接位点的磷酰基团。第二次转酯反应导致两个结果。首先, 当然也是最重要的, 是把5’和3’外显子连接起来了。其次, 使内含子作为离去基团释放出去。
剪接体
RNA剪接是由一个叫做剪接体的大复合体执行的
剪接体(spliceosome): 一个大型分子复合体, 这个复合体包含约150种蛋白质和5种RNA,即使只完成一步剪接反应, 剪接体也需要水解几个分子的ATP。剪接体的多数功能是由其RNA而不是蛋白质组分执行的。
核内小RNA(small nlearRNA, snRNA): 剪接体中的5种RNA(U1、U2、U4、U5和U6)统称为核内小RNA。
小核内核糖蛋白(small nuclear ribonuclear protein, snRNP): 在大多数真核细胞中每种snRNA长100- 300nt,且与几种蛋白质形成复合物, 这些RNA-蛋白质复合物称为小核内核糖蛋白。不同的snRNP在不同时间进出剪接体, 行使特殊的功能, 完成剪接过程。
snRNP在剪接中的功能:
- 识别5剪接位点和分支点;
- 按需要把这两个位点集结到一起;
- 催化(或协助催化)RNA剪切和连接反应。
剪接过程
剪接体内的组装、重排与催化反应:剪接过程
- 首先,5剪接位点被U1 snRNP识别(通过其snRNA与pre -mRNA之间的碱基配对)。U2AF由两个亚单位组成, 较大的一个亚基(65)与多聚嘧啶区结合, 较小的一个亚基(35)与3’剪接位点结合。前一个亚基与BBP(SF1)相互作用, 协助其结合到分支点上。以上形成的蛋白质和RNA结合体称为早期(E)复合体。
- 随后, 在U2AF协助下, U2 snRNP取代BBP(SF1)结合到分支点, 这样形成的结合物称为A复合体。
- 下一步是A复合体通过重排把3个剪接位点拉到一起。这个步骤是通过U4、U6及U5snRNP分步加入复合体而完成的。这3个snRNP合称三联snRNP(tri-snRNP)颗粒, 其中U4和U6 snRNP通过其RNA组分的互补配对相结合, 而U5 snRNP通过蛋白质相互作用松散结合。随着tri-snRNP颗粒的加入, A复合体就转变成B复合体。
- 随后, U1离开剪接体, 其在5’剪接位点的位置由U6取代。这要求U1 snRNA与pre-mRNA之间的碱基配对断开, 以便U6 RNA与同一区域配对(实际上是与一段与之重叠的序列配对)。以上就是全部的组装过程。
- 后续的重排启动了催化反应, 过程如下: U4离开剪接体, 以便U6与U2通过RNA-RNA配对发生作用。重排后的剪接体叫C复合体, 具有活性位点。重排把相关组分(一般认为仅包含U2和U6snRNA)聚集到剪接体内, 从而产生活性位点。
- 活性位点出现后, 就把pre-mRNA的5’剪接位点与分支点拉到了一起, 加速了第一次转酯反应。第一步转酯反应是由分支点保守腺苷酸的2’羟基引发的。它作为亲核基团, 攻击5’剪接位点保守鸟苷酸的磷酰基团。结果, 外显子3’端的核糖与内含子5’端磷酸之间的磷酸二酯键断开, 而游离出来的5’磷酸与分支点保守腺苷酸的2’羟基连接;
- 最后的步骤是释放出mRNA产物及snRNP。在一起。最后的步骤是释放出mRNA产物及snRNP。起初snRNP仍然与内含子形成的套索结合在一起, 随着套索的快速降解, snRNP又进入下一轮循环。
5’和3’剪接位点的第二次转酯反应由U5 snRNP协助, 在第二步转酯反应中, 5’外显子新游离出来的3’羟基反过来作为亲核基团, 攻击3’剪接位点的磷酰基团。第二次转酯反应导致两个结果。首先, 当然也是最重要的, 是把5’和3’外显子连接起来了。其次, 使内含子作为离去基团释放出去。
自剪接内含子的存在表明RNA可以催化RNA剪接
自剪接内含子(self-splicing introns):不需要剪接体,通过自身折叠成一种特殊的构象就可以把自身从pre-mRNA上切下来的内含子。自剪接内含子根据结构和作用机制分为两类:I类(groupI)和I类(groupII)自剪接内含子。
RNA剪接的三种类型
| 类型 | 丰度 | 机制 | 催化机器 |
|---|---|---|---|
| 细胞核pre-mRNA | 很常见, 适用于大多数真核基因 | 两步转酯反应 | 主要剪接体 |
| Ⅱ类自剪接内含子 | 罕见,来自某些细胞器的 | 真核基因及原核基因 | 与 pre-mRNA 相同 内含子编码的核酶 |
| Ⅰ类自剪接内含子 | 罕见,某些真核生物的细胞核 rRNA、细胞器基因,以及少量原核基因 | 两步转酯反应,外源游离G | 与Ⅱ类自剪接内含同 |
对于II类自剪接内含子, 剪接的化学过程以及所产生的RNA中间产物, 与细胞核内pre-mRNA相同。
Ⅰ类自剪接内含子释放出线形内含子,而不是一个套索结构
I类自剪接内含子剪接机制:
它们使用游离的鸟苷或鸟苷酸, 而不是分支点腺苷酸。RNA分子结合该鸟苷或鸟苷酸, 并将其3’羟基呈递给5’剪接位点。以形成套索结构的同类转酯反应, 把游离的鸟苷或鸟苷酸与内含子5’端连接起来。
第二步反应与以前讲述的完全相同: 游离的外显子3’端攻击3’剪接位点。这个反应把两个外显子连接起来并释放出内含子, 尽管在这里内含子是线形的, 而不再是套索状。
I类与II类自剪接内含子的区别:
- Ⅰ类自剪接内含子比Ⅱ类小, 有一个保守的二级结构。该结构包括一个容纳鸟苷或鸟苷酸的结合口袋, 只要这些鸟苷或鸟苷酸是以核糖的形式出现。
- Ⅰ类自剪接内含子还含有一段”内在指导序列”, 与5’剪接位点的序列配对, 因而确定了鸟苷亲核攻击的精确位置。
剪接体怎么可靠地确定剪接位点?
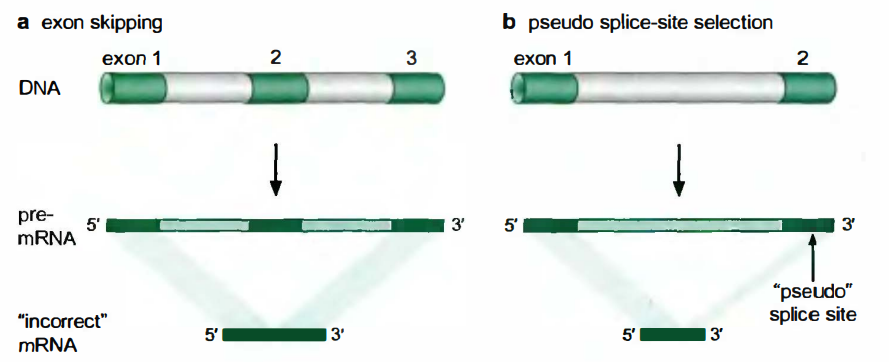
剪接体为什么容易犯错?
- 外显子长度一般只有150nt左右, 而内含子长度平均可达3000nt(前面提到, 有的内含子可长达800000nt)。因此必须从内含子的汪洋大海中捕捞外显子。
- 剪接位点的保守序列相当宽松, 只有AG/G这三个核苷酸对于3’剪接位点是必须的, 这种保守序列在每64nt种就会出现一次。
剪接位点的识别容易犯两种错误:
一是遗漏了剪接位点, 例如, 结合到5’剪接位点的剪接体成分, 跳过了正确的3’剪接位点, 与结合到下游另一个3’剪接位点的剪接体成分配对。
二是与剪接位点邻近的非法位点被错认为剪接位点。
增加剪接位点选择准确性的两种机制:
- 在将某个基因转录为RNA时, RNA复制酶Ⅱ携带了多种具有RNA加工功能的蛋白质, 其中包括参与剪接的蛋白质。当在新合成的RNA分子上遇到5’剪接位点时, 那些蛋白质就从RNA复制酶Ⅱ的C端(剪接相关蛋白质搭乘在RNA复制酶Ⅱ的位置)转移到RNA分子上。剪接体成分一旦结合了5’剪接位点, 就会做好准备与那些结合即将转录出来的、下一个3’剪接位点的剪接体成分相互作用。于是在更下游的任何竞争3’剪接位点被转录出来之前, 剪接体成分就已经识别出了正确的3’剪接位点。
- 确保优先识别邻近外显子的剪接位点(因而更有可能是真正的剪接位点), 以防止使用错误剪接位点。
- SR蛋白结合到外显子中的外显子剪接增强子(ESE)。结合到这些位点的SR蛋白与剪接机器的组分相作用, 将其引导到附近的剪接位点。这样剪接机器就可以更有效地结合到正确的剪接位点, 而不会结合到离外显子较远的错误位点。具体来说, SR蛋白将U2AF蛋白引导到3剪接位点并将U1 snRNP引导到5剪接位点。可通过SR蛋白和剪接体蛋白间的直接相互作用来实现。
- 另外一种方式是通过与剪接体组装和工作时形成的RNA: RNA杂合体发生相互作用及对后者的稳定来实现。
SR蛋白是剪接所必需的。它们不仅保证了基本剪接的准确性和高效性(上面刚刚提到), 而且调节可变剪接。它们种类繁多, 有些由生理信号调控, 另外一些则不受生理调控而具有活性(组成型)。有一些只在某些特定的细胞类型中表达, 调节细胞类型特异性剪接。
剪接替换
不同RNA分子的外显子可通过反式剪接融合
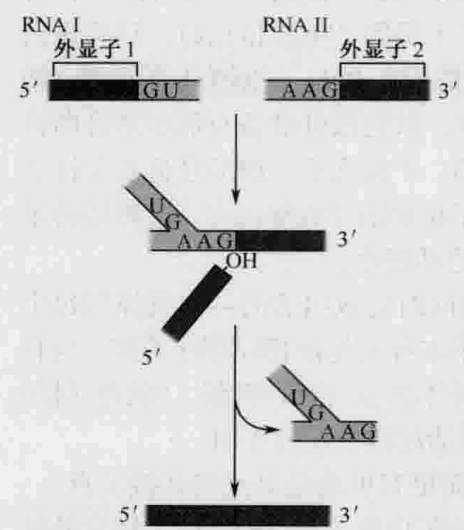
反式剪接(trans-splicing): 把位于在不同pre-RNA分子上的2个外显子连接在一起形成成熟mRNA的过程, 与此同时释放出一个Y形结构的内含子。反式剪接的化学过程与标准剪接方式无异(两步转酯反应), 虽然这种剪接方式在大多数生物中发生的概率很低, 但经常出现在锥虫的mRNA上。
少数内含子是由另一组snRNP组成的另一种剪接体来剪接的
低丰度剪接体识别一类很少见的内含子, 其共有序列不同于绝大多数pre-mRNA内含子的共有序列。这种新发现的低丰度剪接体称为AT-AC剪接体, 因为最初发现其识别的罕见内含子的5’端是AU, 3’端是AC(对应于DNA则为AT和AC)。
可变剪接
通过可变剪接一个基因可以得到多个产物
许多高等真核生物编码的RNA可以通过可变剪接, 产生两条或多条不同的mRNA, 从而翻译出不同的蛋白质。
可变剪接可以组成型的, 也可以是受调控的。在前者, 同一个基因总是产生多种不同的产物;在后者, 不同的产物会在不同的时间、不同的条件, 或在不同的细胞或组织类型中产生。
RNA剪接的5种模式:
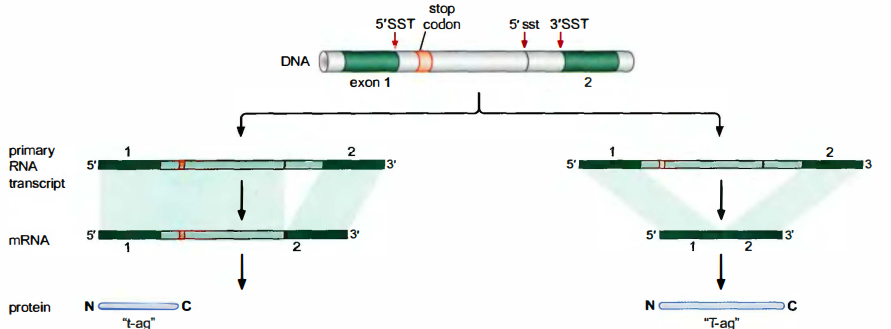
T抗原基因编码两个蛋白质:大T抗原(T-ag)和小t抗原(t-ag)。T抗原基因有2个外显子, 由于使用了不同的5’剪接位点就产生了不同的成熟mRNA。它的mRNA还含有部分内含子序列这个大的mRNA反而编码小蛋白质的原因是在该mRNA所保留的部分内含子序列中有一个终止密码子。
不均一核糖核蛋白(heterogeneous nuclear ribonucleoprotein, hnRNP): 多数减弱子由不均一核糖核蛋白家族的成员识别, 这些蛋白质能结合RNA但没有RS结构域, 所以无法募集剪接体, 相反它们可以阻断特定的剪接位点, 使之丧失作用。
哺乳动物的一个剪接抑制因子是hnRNPI蛋白。有时hnRNPI直接结合到多聚嘧啶区阻断剪接体的结合, 所以也叫多聚嘧啶区结合(Py tract binding, PTB)蛋白。有时它结合到某个外显子两个外侧的序列上, 使该外显子不能进入成熟的mRNA。
可变剪接的结果:
- 产生多个蛋白质产物-异构体, 有相似的, 不同的甚至是相反的功能, 一个基因编码多个功能;
- 开启或关闭只编码一种功能的基因: 外显子含有终止密码子导致产生无功能蛋白, 含有内含子则阻止mRNA转运。
可变剪接受激活因子和抑制因子的调控
剪接调控蛋白结合到称为外显子/内含子剪接增强子(ESS或ISE, exonic/intronic splicingenhancer)或者外显子/内含子剪接减弱子(ESS或ISS, exonic/intronic splicing silencer, ISS)的特殊序列上。前者增强附近的剪接位点的剪接, 后者正好相反。激活因子结合到剪接增强子上, 抑制因子结合到剪接减弱子上。
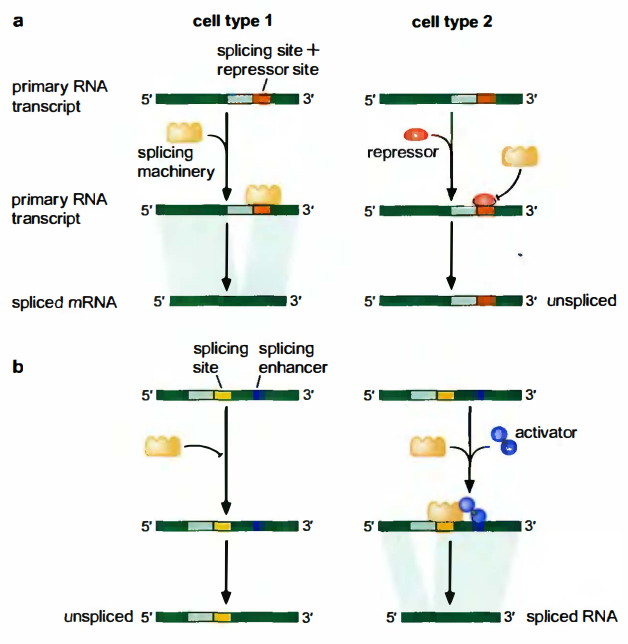
SR蛋白利用某个结构域结合RNA, 如熟知的RNA识别基序(RNA-recognition motif, RRM)。每个SR蛋白都含有另一个结构域, 富含精氨酸和丝氨酸, 称为RS结构域(RS domain)。该结构域位于肽链的C端, 介导SR蛋白与剪接体蛋白的相互作用, 以便把剪接体募集到附近的剪接位点。
外显子重组
外显子通过重组的方式完成改组,从而产生了编码新蛋白质的基因
- 某个特定基因的外显子和内含子的交界处经常与该基因编码蛋白质的结构域的边界相一致, 即每个外显子似乎经常编码蛋白质的一个独立的折叠单位(也经常与一个独立的功能相一致)。
- 许多基因及其所编码的蛋白质明显是在进化过程中部分地通过外显子复制和变异而产生的。
- 有时在其他方面并不相关的基因中会发现相关的外显子, 即有证据显示有些外显子确实在编码不同蛋白质的基因中被重复使用。
RNA编辑
RNA编辑是改变mRNA序列的另一种方法
RNA编辑(RNA editing): 一种可以在RNA转录之后改变其序列, 从而翻译出来的蛋白质与根据基因序列所推导出来的不同的RNA修饰方式。主要有位点特异性脱氨作用和引导RNA指导的尿嘧啶插入或删除两种方式。
RNA编辑的机制: 位点特异性脱氨基作用、引导RNA指导的尿嘧啶插入或删除。
- 位点特异性脱氨基作用(deamination): mRNA中某个特异选择的胞嘧啶经过胞苷脱氨酶脱氨基变成尿嘧啶。对于一种给定的mRNA, RNA编辑只在特定的组织或细胞发生, 而且是受到调控的。其他酶促脱氨的RNA编辑例子包括腺嘌呤的脱氨基作用。这个反应由作用于RNA的腺苷脱氨酶(adenosine deaminaseactingon RNA, ADAR)催化, 产生次黄嘌呤。由于次黄嘌呤可以与胞嘧啶配对, 所以这种编辑形式很容易改变mRNA编码的蛋白质序列。
- 在锥虫线粒体中有另一种截然不同的RNA编辑形式。在这种情况中, RNA转录后, 在pre-mRNA的特定位置插入多个尿嘧啶(有时也可能是删除几个尿嘧啶)。多个尿嘧啶的插入使得mRNA的密码子和可读框发生了改变, 从而完全改变了mRNA的”意思”。尿嘧啶是又引导RNA(guide RNA, gRNA)插入到mRNA中去的。
gRNA结构:
第一区位于5’端, 称为”锚”区, 负责引gRNA到达它所要编辑的mRNA的目标区域;
第二区用于精确定位在被编辑的序列中尿嘧啶将插入的位置;
第三区位于3’端, 是一段多聚尿嘧啶序列。
机制: gRNA的”锚”区有一段序列, 可以与mRNA上将要编辑区域的紧邻位置上(3’端)的一段序列形成碱基配对。随后编辑”指令”发出: gRNA的一段序列与mRNA上的被编辑区互补配对, 但有几个多余的腺嘌呤。这些腺嘌呤在gRNA上的位置与将要在mRNA上插入尿嘧啶的位置正好相反。gRNA与mRNA形成RNA-RNA双链, 而在将要插入尿嘧啶的对应位置形成突出的单链环状结构。一种核酸内切酶识别并切开与环状结构相对的mRNA。由3’端尿苷酰转移酶(TUT酶)催化在mRNA上打开的缺口中加入尿嘧啶。尿嘧啶插入后, 某种RNA连接酶把两段mRNA连接起来, 而gRNA的编辑区则沿mRNA从3’→5’方向继续发挥作用。
RNA编辑的生物学意义:
- 校正作用: 有些基因在突变过程中丢失的遗传信息可以通过RNA编辑得以修复。
- 调控翻译: 通过编辑可以构建或去除起始密码子和终止密码子, 是基因表达调控的一种方式。
- 扩充遗传信息: 能够使基因产物获得新的结构和功能, 有利于生物的进化。
mRNA转运
一旦完成加工,mRNA会包装好并从细胞核输出到细胞质用于翻译
mRNA一旦全部加工完毕(加帽、去除内含子和多聚腺苷酸化)就会运出细胞核进入细胞质, 然后翻译得到蛋白质产物。典型的成熟mRNA携带着一组蛋白质, 正是它们决定了mRNA被转运的命运。
转运出核的条件:
- 成熟mRNA携带残留的SR蛋白
- 携带另外一组专门结合外显子接头的蛋白质(只见于完成剪接的RNA)。
mRNA转运是通过核膜上的一种特殊结构完成的, 这就是核孔复合体(nuclear pore complex)。一旦进入细胞质, 蛋白质组分就会解离下来, 并被识别后重新运回细胞核;然后再与另外的mRNA结合, 重复下一次循环。
损坏的、错误加工的以及剪接释放的内含子都留核内降解。
考点补充
名词补充