细胞生物学第四章物质跨膜运输
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16 __
/\ \
___ ___ __ ___ ___\ \ \____ _ __ __ ___ __
/' __` __`\ /'__`\/' __` __`\ \ '__`\/\`'__\/'__`\ /' _ `\ /'__`\
/\ \/\ \/\ \/\ __//\ \/\ \/\ \ \ \L\ \ \ \//\ \L\.\_/\ \/\ \/\ __/
\ \_\ \_\ \_\ \____\ \_\ \_\ \_\ \_,__/\ \_\\ \__/.\_\ \_\ \_\ \____\
\/_/\/_/\/_/\/____/\/_/\/_/\/_/\/___/ \/_/ \/__/\/_/\/_/\/_/\/____/
__ __
/\ \__ /\ \__
\ \ ,_\ _ __ __ ___ ____ _____ ___ _ __\ \ ,_\
\ \ \/ /\`'__\/'__`\ /' _ `\ /',__\/\ '__`\ / __`\/\`'__\ \ \/
\ \ \_\ \ \//\ \L\.\_/\ \/\ \/\__, `\ \` \L\ \/\ \L\ \ \ \/ \ \ \_
\ \__\\ \_\\ \__/.\_\ \_\ \_\/\____/\ \ ,__/\ \____/\ \_\ \ \__\
\/__/ \/_/ \/__/\/_/\/_/\/_/\/___/ \ \ \/ \/___/ \/_/ \/__/
\ \_\
\/_/
膜转运蛋白与小分子物质的跨膜转运
膜双层的不透性和膜转运蛋白
- 活细胞内外离子浓度不同, 胞外Na+高, 胞内K+高。
- 浓度差的调控机制:
- 膜转运蛋白的活性
- 质膜脂双层的疏水性特征
- 除了脂溶性分子和不带电荷的小分子可以简单扩散的方式进入细胞;脂双层对绝大多数溶质分子和离子是高度不透的, 都需要膜转运蛋白的协助。
- 膜转运蛋白的分类: 载体蛋白和通道蛋白
载体蛋白及其功能
载体蛋白(carrier protein, transporter): 几乎存在所有类型的生物膜上, 属于多次跨膜蛋白。可以介导主动运输和被动运输。每种载体蛋白能与特定的溶质分子结合, 通过一系列构象改变介导溶质分子的跨膜转运。
载体蛋白的特点: 具有高度选择性;有类似于酶的饱和动力学特征;可被竞争性抑制或非竞争抑制;对pH依赖;与酶不同, 载体蛋白对转运的溶质分子不做任何共价修饰;介导主动运输和被动运输。
通道蛋白及其功能
通道蛋白(channel protein):是膜结合蛋白, 通道蛋白形成选择性和门控性跨膜通道, 对离子的选择性取决于通道的直径, 介导被动运输, 通过形成亲水性通道实现对特异溶质的跨膜转运。有3种类型: 离子通道、孔蛋白以及水孔蛋白。大多数通道蛋白都是离子通道。
离子通道的本质是离子选择性通道;依赖通道的直径和形状选择离子;转运底物时, 通道蛋白形成选择性和门控性跨膜通道。
通道蛋白的特点为: 极高的转运速率;由浓度梯度和跨膜电位差(跨膜电化学梯度)来驱动;没有饱和值;具有门控性, 并非持续开放;只介导被动运输。
根据激活信号不同, 离子通道可以分为
- 1.电压门通道(voltage-gated channel):这类通道的构型变化依据细胞内外带电离子的状态, 主要是通过膜电位的变化使其构型发生改变, 从而将”门”打开,一些离子、代谢物或其它溶质顺着浓度梯度自由扩散通过细胞膜。电压门通道在神经细胞的信号传导中起主要作用, 电压门控通道也存在于其他的一些细胞,包括肌细胞、卵细胞、原生动物和植物细胞。
- 2.配体门通道(ligand gated channel): 这类通道在其细胞内或外的特定配体(ligand)与膜受体结合时发生反应, 引起门通道蛋白的一种成分发生构型变化, 结果使”门”打开。因此这类通道被称为配体-门控通道, 它分为细胞内配体和细胞外配体两种类型。
- 3.应力激活通道(stress-activated channel): 通道蛋白感应应力而改变构象, 从而开启通道形成离子流, 产生电信号。听觉毛状细胞的离子通道就是一个极好的例子。
小分子物质的跨膜运输类型
简单扩散
简单扩散(simple diffusion): 简单扩散是指小分子物质以热自由运动的方式顺着电化学梯度或浓度梯度, 不需要细胞提供能量, 也无需膜转运蛋白的协助, 直接通过脂双层进出细胞的运输方式。在简单扩散的跨膜运动中, 脂双层对溶质的通透性大小主要取决于分子大小和分子的极性。疏水性小分子如$O_2$、$N_2$以及小的不带电荷的非极性分子很容易通过简单扩散进出细胞。
过程: 物质先溶解在膜脂中, 再从一侧扩散到另外一侧, 最后进入水相。
物质通透性决定于分子的脂溶性、极性、分子大小和带电性:
- 脂溶性越高, 通透性越强;水溶性越高, 通透性越弱;
- 非极性分子比极性更易透过。$H_2O$、$O_2$等可以透过, 但速度较慢;
- 小分子比大分子更易透过;
- 对带电荷的物质是高度不通透。
被动运输
被动运输(passive transport): 被动运输即协助扩散(facilitated diffusion), 是指溶质顺着电化学梯度或浓度梯度, 不需要细胞提供能量, 在膜转运蛋白协助下的跨膜转运方式。多种极性小分子和无机离子, 包括水分子、糖、氨基酸、核苷酸以及细胞代谢物等顺着电化学梯度或浓度梯度完成跨膜转运。
葡萄糖转运蛋白(glucose transport, GLUT): 具有12次跨膜α螺旋, 通过构象改变完成葡萄糖的协助扩散, 转运方向取决于葡萄糖浓度梯度。
水孔蛋白(aquaporin , AQP): 是水分子的跨膜通道。它是一类具有6个α螺旋区的蛋白质家族, 通常形成四聚体的膜蛋白以行使其转运水或甘油等分子的功能。水分子可以借助质膜上水孔蛋白实现快速跨膜转运, 对水具有特异通透性。它对于细胞渗透压以及生理与病理调节很重要。植物水孔蛋白在种子萌发、细胞伸长、气孔运动以及受精等过程中调节水分的快速跨膜转运。有些水孔蛋白还在植物逆境应答如抗旱性中起着重要作用。
ATP驱动泵与主动运输
主动运输
主动运输(active transport): 是由载体蛋白所介导的物质逆着电化学梯度或浓度梯度进行跨膜转运的方式, 需要消耗能量。主动运输普遍存在于动、植物细胞和微生物细胞。根据能量的来源不同, 可将主动运输分为: ATP直接提供能量、间接提供能量以及光驱动泵3种基本类型。
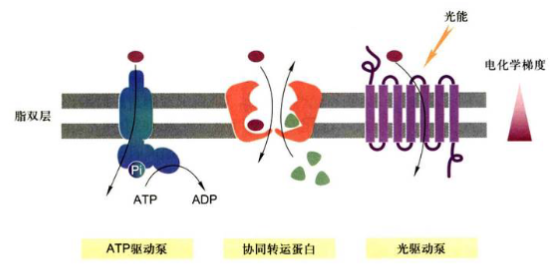
- 1.ATP驱动泵(ATP-driven pump): 是ATP酶直接利用水解ATP提供能量, 将ATP水解生成ADP和Pi, 实现离子或小分子逆浓度梯度或电化学梯度的跨膜运输。这一类主动运输是一种能量偶联的化学反应过程, 即离子或小分子逆化学梯度的”上山”运动和ATP水解(释放能量)相偶联。
- 2.协同转运蛋白(cotransporter): 又称为偶联转运蛋白(coupled transporter)介导各种离子和分子的跨膜运动。这一类蛋白包括两种基本类型: 同向协同转运蛋白(symporter)和反向协同转运蛋白(antiporter)。这两类转运蛋白可以使一种离子或分子逆浓度梯度的转运与另一种或多种其他溶质顺着电化学梯度或浓度梯度偶联。区别于ATP驱动泵直接供能, 协同转运蛋白利用的能量储存在其中一种溶质的电化学梯度中。
- 1.同向协同转运(symport): 是偶联物的运输方向相同, 如小肠上皮细胞吸收葡萄糖伴随着Na+从细胞外流入细胞内而完成的;
- 2.反向协同转运(Antiport): 是偶联物的运输方向相反, 如质膜上H+/Na+交换载体在完成H+输出细胞的同时伴随着Na+输入细胞。
例如, 在动物细胞的质膜上, Na+是常用的协同转运离子, 它的电化学梯度为另一种分子的主动运输提供了驱动力, 由于Na+电化学梯度的形成需要Na+-K+泵水解ATP, 事实上, 协同转运是一种间接消耗主动转运的方式。
在细菌、酵母和动物细胞的被膜细胞器, 绝大多数协同运输是靠H+而不是靠Na+电化学梯度来驱动的。
- 3.光驱动泵(light-driven pump): 主要发现于细菌细胞, 对于溶质的主动运输与光能的输入相偶联, 如菌紫红质。
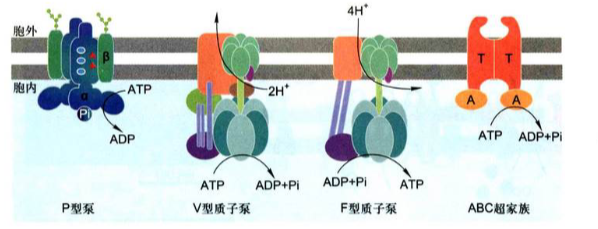
P型泵
P型泵(P-type pump): 有两个独立的α催化亚基, 具有ATP结合位点, 绝大多数还具有两个起调节作用的小的β亚基。在转运离子过程中, 至少有一个α催化亚基发生磷酸化和去磷酸化反应, 改变转运泵的构象, 实现离子的跨膜转运。由于转运泵水解ATP使自身形成磷酸化中间体, 因此称作P型泵。包括$Na^+$-$K^+$泵、$Ca^{2+}$离子泵和$H^+$离子泵。
$Na^+$-$K^+$泵
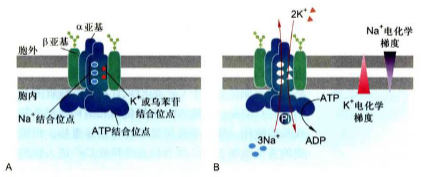
结构特点
Na+-K+泵(Na+-K+ pump): 又称为Na+-K+ ATPase, 是位于动物细胞的质膜上的载体蛋白, 具有ATP酶活性, 在ATP直接功能的条件下能逆浓度梯度主动转运钠离子和钾离子。由2个α和β亚基组成的四聚体, 工作时, 通过α亚基一个天冬氨酸残基的磷酸化和去磷酸化使α亚基的构象改变来实现钠钾的排出和吸入。
转运机制
细胞内侧a亚基与Na+相结合促进ATP水解, a亚基上的一个天冬氨酸残基磷酸化引起a亚基构象发生变化, 将Na+泵出细胞, 同时细胞外的K+与a亚基的另一位点结合, 使其去磷酸化, a亚基构象再度发生变化将K+泵入细胞, 完成整个循环。Na+依赖性的磷酸化和K+依赖型的去磷酸化引起Na+-K+泵构象发生有序变化, 每次循环消耗一个ATP分子, 可以逆着电化学梯度进行跨膜转运。
乌本苷(ouabain): 极少量便可抑制Na+-K+泵的活性, 它通过结合K+结合位点来作用。此外, 生物氧化抑制剂如氰化物使ATP供应中断, Na+-K+泵失去能源以致停止工作。而Mg+和少量膜脂有助于Na+-K+泵活性的提高。
生理功能
- 维持细胞膜电位: 细胞质膜两侧具有一定的电位差, 即膜电位, 每一个工作循环下来, Na+-K+泵将从细胞泵出3个Na+泵入2个K+, 对膜电位的形成起到了一定作用。
- 维持动物细胞渗透的平衡: 如果没有Na+-K+泵将Na+泵出细胞, 水分子将由于渗透压的缘故顺着自身的浓度通过水孔蛋白大量进入细胞引起细胞吸水膨胀。(与动物不同, 植物细胞依靠坚韧的细胞壁防止膨胀和破裂, 能耐受较大的跨膜渗透差异。
- 吸收营养: 动物细胞对葡萄糖或氨基酸等有机物吸收的能量由蕴藏在Na+电化学梯度中的势能提供。动物细胞利用膜两侧的Na+电化学梯度以协同转运的方式吸收营养物, 而植物细胞、真菌和细菌通常利用质膜上的H+-ATPase形成的H+电化学梯度来吸收营养物。
$Ca^{2+}$离子泵
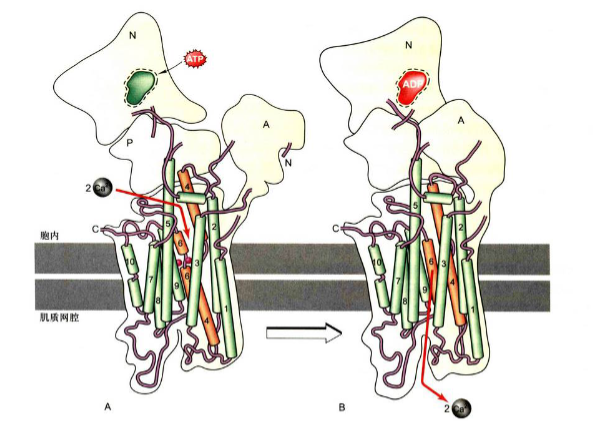
结构特点
Ca2+泵(Ca2+ pump): 又称为Ca2+-ATPase, 是一类P型泵, 是由1000个氨基酸组成的跨膜蛋白, 主要分布在质膜或细胞器膜上。可以将Ca2+泵到细胞外或内质网腔内从而维持细胞质基质中低Ca2+浓度。对于肌肉细胞来说, Ca2+泵将Ca2+泵入肌质网, 对调节肌细胞的收缩运动至关重要。
转运机制
原理与钠钾泵相似, 每分解一个ATP, 泵出2个Ca2+, 将Ca2+输出细胞或泵入内质网腔中储存起来。
P型H+泵
结构特点
H+泵(H+ pump): P型H+泵将H+泵出细胞, 建立和维持跨膜的H+电化学梯度, 并用来驱动转运溶质进入细胞。植物细胞、真菌(包括酵母)和细菌细胞质膜上有P型H+泵。
转运机制
利用ATP自磷酸化导致的构象改变来转移H+。如植物细胞膜上的H+泵、动物胃表皮细胞的H+-K+泵(分泌胃酸)。
V型质子泵和F型质子泵
V型质子泵(V-type proton pump): 位于动物细胞胞内体膜、溶酶体膜, 破骨细胞和某些肾小管细胞的质膜, 以及植物、酵母及其他真菌细胞的液泡膜上, 由许多跨膜和胞质亚基构成, 水解ATP产生能量, 将H+泵入细胞器, 以维持细胞质基质pH中性和细胞器内pH酸性, 但不形成磷酸化中间体。
F型质子泵(F-type proton pump): 存在于细菌质膜、线粒体内膜和叶绿体类囊体膜上, 含有几种不同的跨膜和胞质侧亚基。利用质子动力势合成ATP, 即当H+顺着电化学梯度通过质子泵时, 所释放的能量驱动F型质子泵合成ATP, 并且在转运H+是不形成磷酸化中间体。因此F型质子泵又被称为H+-ATP合成酶。
ABC超家族
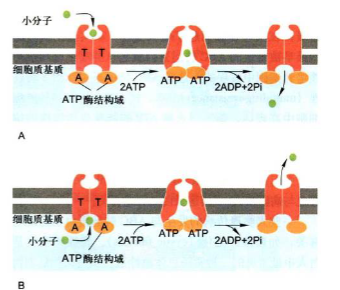
结构
ABC超家族(ABC superfamily): 是一类ATP驱动泵, 又叫ABC转运蛋白(ATP-binding cassette)。该超家族含有几百种不同的转运蛋白, 广泛分布于细菌到人各种生物中, 是最大的一类转运蛋白。每种ABC转运蛋白对于底物或底物的集团有特异性, 都由4个”核心”结构域组成。ABC转运蛋白能将天然毒物和代谢废物排出体外, 也能够将抗生素或其他抗癌药物泵出细胞而赋予细胞抗药性。
工作模式
- ATP分子与ABC转运蛋白结合, 诱导ABC转运蛋白2个ATP结合域二聚化, 引起转运蛋白构象改变, 使底物结合部位暴露于质膜的另一侧;
- ATP水解以及ADP的解离将导致ATP结合域解离, 引起转运蛋白构象恢复原有状态。
ABC转运蛋白与疾病
ABC蛋白能够将抗生素或其他抗癌药物泵出细胞而赋予细胞抗药性。
多药抗性转运蛋白(multidrug-resistance, MDR): 属于一种ABC转运蛋白, 在多种肿瘤细胞中高表达, 能利用水解ATP的能量将脂溶性的抗癌药物从细胞内转运到细胞外, 从而降低细胞内药物浓度, 导致肿瘤细胞抗药性增强而降低患者化疗效果。
离子跨膜转运与膜电位
胞吞作用与胞吐作用
膜泡运输(vesicular transport): 又称为批量运输(bulk transport)。细胞对大分子及颗粒性物质的跨膜转运方式。包括内吞作用、外吐作用两个不同方向的物质转运过程。这个需要ATP供能的运输活动涉及细胞膜或胞内膜的变形、膜性水泡的形成与膜泡融合等过程, 被转运的物质包裹在脂双层膜围成的囊泡中, 故称膜泡运输。它是蛋白质分选运输途径之一, 蛋白质被选择性地包装成运输小泡, 定向转运到靶细胞器。
胞吞作用(endocytosis): 细胞通过质膜内陷形成囊泡, 将胞外的生物大分子、颗粒性物质或液体等摄取到细胞内, 以维持细胞正常的代谢活动。根据胞吞泡形成的分子机制不同和胞吞泡大小的差异分为胞饮作用和吞噬作用。
胞吐作用(excytosis): 细胞内合成的生物分子(蛋白质和脂质等)和代谢物以分泌泡的形式与质膜融合将内含物分泌到细胞表面或细胞外的过程。主要分为组成型胞吐途径和调节型胞吐途径。其中组成型胞吐途径通过新合成的蛋白质和脂质以囊泡形式连续不断地供应质膜更新, 确保细胞分裂前质膜的生长。调节型胞吐途径是特化的分泌细胞产生的分泌物储存在分泌泡内, 当细胞在受到胞外信号刺激时, 分泌泡与质膜融合并将内含物释放出去。
胞吞泡(endocytic vesicle): 胞吞时质膜内陷脱落形成的囊泡, 称胞吞泡。
胞吞作用的类型
吞噬作用
吞噬作用(phagocytosis): 细胞内吞较大的固体颗粒物质, 称为胞吞作用。吞噬作用是原生生物摄取食物的一种方式。在高等多细胞生物体中, 吞噬作用往往发生于巨噬细胞和中性粒细胞, 其作用不仅仅是摄取营养物, 主要是清除侵染机体的病原体以及衰老或凋亡的细胞。
胞饮作用
胞饮作用(pinocytosis): 细胞对液体物质或细微颗粒物质的摄入和消化过程。当细胞对这类物质进行转运时, 由质膜内陷形成胞饮泡, 将待转运的物质包裹起来进入细胞质。胞饮作用可以分为网格蛋白依赖的胞吞作用、胞膜窖依赖的胞吞作用、大型胞饮作用以及非网格蛋白/胞膜窖依赖的胞吞作用。大多数真核细胞都能通过胞饮作用连续摄入溶液及可溶性分子。
网格蛋白依赖的胞吞作用(clathrin dependent endocytosis)
网格蛋白(clathrin): 是一种进化上高度保守的蛋白质, 由3个二聚体组成, 每个二聚体包括1条重链和1条轻链。3个二聚体形成三脚蛋白复合体(triskelion), 是包被的结构单位。
网格蛋白包被膜泡形成机制:
当配体与膜上受体结合后, 网格蛋白聚集在膜下, 逐渐形成直径50~100nm的质膜凹陷, 称网格蛋白包被小窝。一种小分子GTP结合蛋白一发动蛋白(dynamin)在深陷的包被小窝的颈部组装成环, 发动蛋白水解与其结合的GTP引起颈部缢缩, 最终脱离质膜形成网格蛋白包被膜泡(clathrin-coated vesicle)。几秒钟后, 网格蛋白便脱离包被膜泡返回质膜附近区域以便重复使用, 脱包被的囊泡与早胞内体(earlyendosome)融合, 从而将转运分子及胞外液体摄入细胞。
需要注意的是, 在大分子跨膜转运中, 网格蛋白本身并不起捕获特异转运分子的作用, 有特异性选择作用的是包被中另一类衔接蛋白(adaptin), 它既能结合网格蛋白, 又能识别跨膜受体胞质面的尾部肽信号, 从而通过网格蛋白包被膜泡介导跨膜受体及其结合配体的选择性运输。
根据胞吞的物质是都具有专一性, 可将网格蛋白介导胞吞作用分为受体介导的胞吞作用和非特异性的胞吞作用。
受体介导的胞吞作用(receptor mediated endocytosis):
绝大多数动物细胞从胞外摄取特定大分子的有效途径, 是一种选择性浓缩机制, 保证了摄入大量特定的分子, 而避免了摄入细胞外大量的液体。如低密度脂蛋白的摄入就是通过这一途径进行的。
重要的例子包括动物细胞通过受体介导的胞吞作用对胆固醇的摄取、鸟类卵细胞摄取卵黄蛋白以及肝细胞摄入转铁蛋白等。某些激素如胰岛素与靶细胞表面受体结合进入细胞, 巨噬细胞通过表面受体对免疫球蛋白及其复合物、病毒、细菌乃至衰老细胞的识别和摄入, 以及其他一些基本代谢物如合成血红蛋白所必需的维生素B12和铁的摄取都是通过受体介导的胞吞作用进行的。受体介导的胞吞作用也可以被某些病毒所利用, 流感病毒和AIDS病病毒(HIV)就是通过这种胞吞途径侵染细胞的。
胆固醇运输步骤:
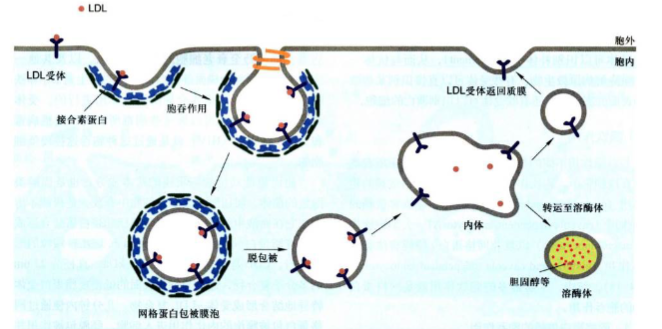
胆固醇是动物细胞质膜的基本成分, 也是固醇类激素的前体。胆固醇主要在肝细胞中合成, 是极端不溶的, 它在血液中的运输是通过与磷脂和蛋白质结合形成低密度脂蛋白颗粒的形式进行。
低密度脂蛋白(low-density lipoprotein, LDL): LDL是相对分子质量为3x10^6, 直径为22 nm的多分子复合物, 通过与细胞表面的低密度脂蛋白受体特异地结合形成受体-LDL复合物, 几分钟内便通过网格蛋白包被膜泡的内化作用进人细胞, 经脱包被作用并与胞内体融合。
胞内体(endosome): 胞内体是动物细胞内由膜包裹的细胞器, 其作用是传输由胞吞作用新摄入的物质包裹的细胞器, 其作用是传输由胞吞作用新摄入的物质到溶酶体。胞内体膜上有ATP驱动的质子泵, 将H+泵入胞内体腔中, 使腔内的pH降低(pH 5~6)。
在此过程中, 低pH环境可引起LDL与受体分离, 而胞内体以出芽的方式形成含有受体的小囊泡, 返回细胞质膜, 受体重复使用。然后含有LDL的胞内体与溶酶体融合,低密度脂蛋白被水解, 释放出胆固醇和脂肪酸供细胞利用。在胞吞过程中, 胞内体被认为是膜泡运输的主要分选站之一, 其中的酸性环境在分选过程中起关键作用。
不同类型的受体具有不同的胞内体分选途径:
- 大部分受体返回它们原来的质膜区域, 如上述LDL受体循环到质膜再利用;
- 受体下行调节(receptor down-regulation): 有些受体不能再循环而是最后进入溶酶体被消化, 如与表皮生长因子(epidermal growth factor, EGF)结合的细胞表面受体,大部分在溶酶体被降解, 从而导致细胞表面EGF受体浓度降低, 这种现象称为受体下行调节。
- 转胞吞作用(transcytosis): 转胞吞作用是一种特殊的内吞作用, 受体和配体在内吞中并未作任何处理, 通过内吞作用被摄入细胞的一侧, 再以外排作用从另一侧输出。这种内吞作用主要发生在极性细胞中, 如抗体转运到血液和奶汁就是这种运输。
其他类型的胞饮作用
- 胞膜窖依赖的胞吞作用: 胞膜窖在质膜的脂筏区域形成, 电镜观察发现有些细胞的胞膜窖呈内陷的瓶状。
- 胞膜窖的特征性蛋白是窖蛋白。胞膜窖的形成部位位于质膜的脂筏区域。
- 形成机制: 胞吞时, 胞膜窖携带着内吞物, 利用发动蛋白的收缩作用从质膜上脱落, 然后转交给胞内体样的细胞器——膜窖体(caveosome)或者跨细胞转运到质膜的另一侧。在整个过程中, 窖蛋白始终不会从胞吞泡膜上解离下来。由于胞膜窖所在部位含有大量信号转导的受体和蛋白激酶等, 这暗示胞膜窖很可能发挥了一种细胞信号转导的平台作用。
- 大型胞饮作用: 是另一种胞饮作用, 它是通过质膜皱褶包裹内吞物形成囊泡完成胞吞作用。
- 与吞噬作用类似之处: 大型胞饮作用形成的胞吞泡也比较大, 质膜皱褶的形成过程也依赖微丝及其结合蛋白。
- 与吞噬作用的区别: 启动吞噬作用的受体往往位于特异细胞表面, 而启动大型胞饮作用的受体却位于很多类型的细胞表面, 受体还能启动其他生理功能, 如有些受体就是与细胞生长相关的生长因子。
- 非网格蛋白/胞膜窖依赖的胞吞作用, 如位于淋巴细胞膜上的白介素2(interleukin-2,IL-2)受体就是介导非网格蛋白/胞膜窖依赖的胞吞作用。
胞吞作用与细胞信号转导
- 1.胞吞作用对信号转到的下调
表皮生长因子及其受体的胞吞作用: EGF是一类相对分子质量较小的胞外信号蛋白分子, 能够刺激上皮细胞及其他多种细胞增殖。与LDL受体不同, 当EGF受体与其配体EGF结合后, 受体二聚化并引起受体胞质结构域酪氨酸残基自磷酸化而被活化, 引起细胞下游信号级联反应;另一方面, 细胞通过胞吞作用, 将EGF受体及EGF吞人细胞内降解, 从而导致细胞信号转导活性下调。这种调节作用即受体下行调节。
- 2.胞吞作用对信号转导的激活:(Notch信号通路)
胞吐作用
组成型胞吐途径(constitutive exocytosis pathway): 在真核细胞, 有高尔基体反面囊膜分泌的囊泡向质膜流动并与之融合的膜泡运输过程, 呈连续分泌状态, 完成质膜更新, 分泌胞外基质组分、营养或信号分子等功能。
调节型胞吐作用(regulated exocytosis pathway): 在真核生物的一些特化细胞, 所产生的分泌物储存在分泌泡内, 当细胞受到胞外刺激时, 分泌泡与质膜合并并将内含物分泌出细胞。该胞吐作用方式称为调节型胞吐途径。
考点补充
请说明Ca2+、CO2、乙醇、葡萄糖、水等五种物质分别是通过何种运输方式通过质膜的?各运输过程是否需要消耗能量?(2010)
- 钙离子(Ca²+)
- 运输方式: Ca²+可以通过多种方式通过细胞膜, 包括Ca²+泵、Ca²+通道
- Ca²+泵进行的主动运输需要消耗ATP能量。
- Ca²+通道进行的被动运输不需要消耗能量, 它允许Ca+顺浓度梯度流动。IP3门控Ca2+通道
- 二氧化碳(CO2)
- 运输方式: CO2通过自由扩散(简单扩散)穿过细胞膜。
- 是否消耗能量: 自由扩散不需要消耗能量, 因为CO2分子顺着浓度梯度移动。
- 乙醇
- 运输方式: 乙醇也是通过自由扩散穿过细胞膜。根据有机物相似相容原理, 乙醇等有机物可以自由扩散通过磷
- 脂双分子层而不受阻碍, 同样的还有乙二醇、甘油(丙三醇)、苯等。
- 是否消耗能量: 自由扩散不需要消耗能量。
- 葡萄糖
- 运输方式: 葡萄糖的运输依赖于具体的环境条件。在红细胞中, 葡萄糖通过协助扩散, 借助GLUT1(一种葡萄糖转运蛋白)进行。在小肠上皮细胞和其他许多细胞中, 葡萄糖通过协同运输(例如, 与Na+一起通过SGLT1)进入细胞, 然后通过协助扩散(如通过GLUT2)离开细胞。
- 是否消耗能量:
- 协助扩散不需要消耗能量。
- 协同运输是一种主动运输, 需要消耗能量。
- 水
- 运输方式: 水分子主要通过水通道蛋白(aquaporins)进行快速转运, 这是一种特殊的协助扩散形式。此外, 在没有水通道的情况下, 水分子也能通过脂双层进行自由扩散, 尽管速度较慢。
- 是否消耗能量: 无论是通过水通道还是自由扩散, 水分子的运输都不需要消耗能量。
小肠上皮细胞膜上载体蛋白转运葡萄糖为何有时是协助扩散, 有时是协同运输?
(2003考点是关于小肠上皮细胞游离面和基底面转运葡萄糖的方式不同来出题的.答题思路.先简单描述小肠上皮细胞的结构: 然后分别描述结构中的不同部位是通过如何方式来吸收葡萄糖的: 紧接着叙述一下为什么同样是小肠上皮细胞, 每个部分吸收葡萄糖的方式会不同?最后总结一下。
小肠上皮细胞是极性细胞, 有面向肠腔的顶面或游离面, 以及基底面。游离面质膜与基底面质膜担负不同的功能, 游离面含有大量吸收葡萄糖分子的协同转运载体, 完成Na+驱动的葡萄糖同向转运;而基底面含有执行被动运输的葡萄糖转运载体, 将葡萄糖转运到细胞外液, 从而完成葡萄糖的吸收和转运功能。由于紧密连接限制了膜蛋白和膜脂分子的流动性, 使得上皮细胞游离面与基底面的膜蛋白以及膜脂分子只能够在各自的膜区域流动, 以行使各自不同的功能。
- 1.协同运输: 当小肠中的葡萄糖浓度较低时, 或者为了对抗浓度梯度将葡萄糖从低浓度区域(如小肠腔)转运到高浓度区域(如细胞内部), 细胞会使用协同运输。这种情况下, 葡萄糖的转运是通过Na+/葡萄糖协同转运蛋白(SGLT, sodium-glucose linked transporter)实现的。SGLT通常位于小肠上皮细胞的顶端膜(apical membrane), 即面向小肠腔的那一侧。SGLT能够结合Na+和葡萄糖, 利用Na+从高浓度(细胞外)向低浓度(细胞内)移动时所产生的能量梯度, 同时将葡萄糖带入细胞内, 即使葡萄糖的浓度梯度是逆向的。Na+/K+ ATP酶(钠钾泵)在细胞侧面(basolateral membrane)不断工作, 将Na?泵出细胞, 从而保持Na?的电化学梯度, 为SGLT提供能量。
- 2.协助扩散: 当细胞内的葡萄糖需要被转运到血液中时, 或者当小肠腔中的葡萄糖浓度远高于细胞内时, 细胞会使用协助扩散。这种情况下, 葡萄糖通过位于小肠上皮细胞基底侧膜(basolateral membrane)的葡萄糖载体蛋白(如GLUT2, glucose transporter type 2)进行转运。GLUT2等载体蛋白帮助葡萄糖顺浓度梯度移动, 不需要额外的能量输入, 因为葡萄糖是从高浓度区域向低浓度区域扩散。
简述真核细胞中质子泵的种类、存在部位和功能
以动物细胞从胞外选择性摄取低密度脂蛋白LDL为例, 说明受体介导的网络(或称笼形)蛋白有被小泡内吞过程及生理意义。(2002)
名词补充
receptor mediated endocytosis
receptor mediated endocytosis受体介导的内吞作用是指细胞依靠细胞表面的受体特异性地摄取细胞外蛋白或其他化合物的过程, 是绝大多数动物细胞从胞外摄取特定大分子的有效途径, 是一种选择性浓缩机制, 其具有高度特异性, 保证了摄入大量特定的分子, 而避免了摄入细胞外大量的液体。如细胞摄取胆固醇过程就是受体介导的内吞作用。
excytosis
胞吐作用(excytosis): 细胞内合成的生物分子(蛋白质和脂质等)和代谢物以分泌泡的形式与质膜駐合将内含物分泌到细胞表而或细胞外的过程。主要分为组成型胞吐途径和调节型胞叶途径。其中组成型胞吐途径通过新合成的蛋白质和脂质以囊泡形式连续不断地供应质膜更新, 确保细胞分裂前质膜的生长。调节型胞吐途径是特化的分泌细胞产生的分泌物储存在分泌泡内, 当细胞在受到胞外信号刺激时, 分泌泡与质膜融合并将内含物释放出去。
channel protein
通道蛋白(channel protein): 是膜结合蛋白, 通道蛋白形成选择性和门控性跨膜通道, 对离子的选择性取决于通道的直径, 介导被动运输, 通过形成亲水性通道实现对特异溶质的跨膜转运。有3种类型: 离子通道、孔蛋白以及水孔蛋白。大多数通道蛋白都是离子通道。
carrier protein , transporter
载体蛋白(carrier protein, transporter): 几乎存在所有类型的生物膜上, 属于多次跨膜蛋白。可以介导主动运输和被动运输。每种载体蛋白能与特定的溶质分子结合, 通过一系列构象改变介导溶质分子的跨膜转运。
passive transport
被动运输(passive transport): 被动运输即协助扩散(facilitated diffusion), 是指各种极性小分子和无机离子如糖、氨基酸、核苷酸以及细胞代谢产物等顺浓度梯度或电化学梯度的跨膜运输, 该过程不需要细胞提供能量, 但需要特异性的膜转运蛋白协助, 从而加快转运速度和提高特异性。
endocytosis
胞吞作用(endocytosis): 细胞通过质膜内陷形成囊泡, 将胞外的生物大分子、颗粒性物质或液体等摄取到细胞内, 以维持细胞正常的代谢活动。根据胞吞泡形成的分子机制不同和胞吞泡大小的差异分为胞饮作用和吞噬作用。
active transport
主动运输(active transport): 是由载体蛋白所介导的物质逆着电化学梯度或浓度梯度进行跨膜转运的方式, 需要消耗能量。主动运输普遍存在于动、植物细胞和微生物细胞。根据能量的来源不同, 可将主动运输分为: ATP直接提供能量、间接提供能量以及光驱动泵3中基本类型。
ion channel
离子通道(ion channel): 是通道蛋白的类型之一, 能够介导被动运输, 通过形成亲水性通道实现对特异溶质的跨膜转运。离子通道可以分为电压门通道、配体门通道和应力激活通道。
Transcytosis
转胞吞作用(Transcytosis)。转胞吞作用是一种特殊的内吞作用, 受体和配体在内吞中并未作任何处理, 通过内吞作用被摄入细胞的一侧, 再以外排作用从另一侧输出。这种内吞作用主要发生在极性细胞中, 如抗体转运到血液和奶汁就是这种运输。
aquaporin, AQP
水孔蛋白(aquaporin, AQP)是水分子的跨膜通道。它是一类具有6个$\alpha$螺旋区的蛋白质家族, 通常形成四聚体的膜蛋白以行使其转运水或甘油等分子的功能。水分子可以借助质膜上水孔蛋白实现快速跨膜转运, 对水具有特异通透性: 它对于细胞渗透压以及生理与病理调节很重要.植物水孔蛋白在种子萌发、细胞伸长、气孔运动以及受精等过程中调节水分的快速跨膜转运: 有些水孔蛋白还在植物逆境应答如抗旱性中起着重要作用
vesicular transport
膜泡运输(vesicular transport): 又称为批量运输(bulk transport)。细胞对大分子及颥粒性物质的跨膜转运方式: 包括内吞作用、外叶作用两个不同方向的物质转运过程.这个需要ATP供能的运输活动涉及细胞膜或胞内膜的变形、膜性水泡的形成与膜泡融合等过程, 被转运的物质包裹在脂双层膜围成的囊泡中, 故称膜泡运输。它是蛋白质分选运输途径之一蛋白质被选择性地包装成运输小泡, 定向转运到靶细胞器:
clathrin
网格蛋白(clathrin): 是一种进化上高度保守的蛋白质, 由3个二体组成, 每个二聚体包括1条重链和1条轻链: 3个二聚体形成三脚蛋白复合体(triske№n), 是包被的结构单位: 02 +泵a2 pump): 又称为Ca2*-ATPase , 是一类p型泉, 是由1 0个氨基酸组成的跨膜蛋白, 主要分布在质膜或细胞器膜上: 可以将ca2+ 泵到细胞外或内质网腔内从而维持细胞质基质中低ca2 +浓度.对于肌肉细胞来说, ca2 +泵将ca2 +泵入肌质网, 对调节肌细胞的收缩运动至关重要:
endocytic vesicle
胞吞泡(endocytic vesicle): 胞吞时质膜内陷脱落形成的囊泡, 称胞吞泡.根据胞吞泡形成的分子机制不同和胞吞泡大小的差异, 胞吞作用可分为两种类型: 吞噬作用和胞饮作用。配体门通道(ligand gated channel): 配体门离子通道是指仅在细胞外的配体(如乙酰胆碱等化学信号)与细胞表面的受体结合时发生反应, 引起通道蛋白构象发生改变, 使闸门开放的离子通道.配体门离子通道属于离子通道:
voltage-gated channel
电压门通道(voltage-gated channel): 属于离子通道的一种类型, 这类通道的构型变化依据细胞内外带电离子的状态, 主要是通过膜电位的变化使其构型发生改变, 从而将”门”打死一些离子、代谢物或其它溶质顺着浓度梯度自由扩散通过细胞膜: 电压门通道在神经细胞的信号传导中起主要作用, 电压门控通道也存在于其他的一些细胞, 包括肌细胞、卵细胞、原生动物和植物细胞。
ABC superfamily
ABC超家族(ABC superfamily): ABC超家族是一类ATP驱动的膜转运蛋白, 利用ATP水解释放的能量将多肽及多种小分子物质进行跨膜转运.ABC超家族包含有几百种不同的转运蛋白, 广泛分布于从细菌到人类的各种生物中, 所有ABC蛋白一般含有4个核心结构域.两个跨膜结构域(T), 形成运输分子的跨膜通道: 两个胞质侧ATP结合域(A), 具有ATP酶活性.
cotransport
协同运输(cotransport): 协同运输又称偶联主动运输,它不直接消耗ATP, 但要间接利用自由能,并且也是逆浓度梯度的运输。运输时需要先建立电化学梯度, 在动物细胞主要是靠钠泵, 在植物细胞则是由H+泵建立的H+质子梯度。
作业
比较载体蛋白与通道蛋白的异同。
比较P型离子泵、V型质子泵、F型质子泵和ABC超家族的异同。
说明Na+-K+泵的工作原理及其生物学意义。
试述胞吞作用的类型与功能。