分子生物学 第四章 DNA的结构
1
2
3
4
5
6
7 ____ __ __ ___ ____ __ __ ______
/\ _`\ /\ \__ /\ \__ /'___\ /\ _`\ /\ \/\ \/\ _ \
\ \,\L\_\ \ ,_\ _ __ __ __ ___\ \ ,_\ __ __ _ __ __ ___ /\ \__/ \ \ \/\ \ \ `\\ \ \ \L\ \
\/_\__ \\ \ \/ /\`'__\/\ \/\ \ /'___\ \ \/ /\ \/\ \/\`'__\/'__`\ / __`\ \ ,__\ \ \ \ \ \ \ , ` \ \ __ \
/\ \L\ \ \ \_\ \ \/ \ \ \_\ \/\ \__/\ \ \_\ \ \_\ \ \ \//\ __/ /\ \L\ \ \ \_/ \ \ \_\ \ \ \`\ \ \ \/\ \
\ `\____\ \__\\ \_\ \ \____/\ \____\\ \__\\ \____/\ \_\\ \____\ \ \____/\ \_\ \ \____/\ \_\ \_\ \_\ \_\
\/_____/\/__/ \/_/ \/___/ \/____/ \/__/ \/___/ \/_/ \/____/ \/___/ \/_/ \/___/ \/_/\/_/\/_/\/_/
DNA由多核苷酸链构成
DNA的基本结构
2’-脱氧核糖与碱基通过糖苷键相连形成脱氧核苷,进而与磷酸通过磷酸酯键相连形成脱氧核苷酸。个核苷酸2’-脱氧核糖上的3’-OH与另一个核苷酸上5’-OH的磷酸基团以磷酸二酯键相连形成了规则、不断重复地多核苷酸链,磷酸二酯键赋予了DNA链的固有极性,DNA链的一端是游离的5’-磷酸基或5’-羟基,另一端是游离的3’磷酸基或3’-羟基。
多核苷酸链中碱基排列顺序的不规则性再加上其长度是DNA储存大量信息的基础。
DNA最重要的结构特征是两条多核苷酸链以双螺旋形式相互缠绕。双螺旋两条单链骨架由糖和磷酸基团交替构成,碱基朝骨架内侧。
DNA的高级结构
- DNA的一级结构:指4种核苷酸的连接及其排列顺序,表示了该DNA分子的构成,每个DNA分子所具有的特定碱基排列顺序构成了DNA分子的特异性。
- DNA的二级结构:指两条多核苷酸链反向平行盘绕所生成的双螺旋结构,DNA的二级结构分两大类:一类是右手螺旋,如A-DNA和B-DNA,另一类是左手螺旋,如Z-DNA。
- DNA的三级结构:指DNA双螺旋进一步扭曲螺旋所形成的更复杂的特定的空间结构。其中超螺旋是主要的形式。
- DNA的四级结构:是指由组蛋白、非组蛋白与DNA组装形成的复杂空间结构,即染色体结构。
每个碱基都有它偏好的异构体
构成DNA的碱基可以分为两类:嘌呤和嘧啶,嘌呤为腺嘌呤和鸟嘌呤;嘧啶为胞嘧啶和胸腺嘧啶。胞嘧啶上的N原子通常以氨基存在,很少以亚氨基存在;鸟嘌呤上的氧原子通常以酮式存在,很少以烯醇式存在。
异构体是碱基在氢键供体与氢键受体之间的转换,是导致DNA合成出错的普遍原因。
双螺旋的两条链通过碱基配对反向平行方式连接
反向平行是A与T,C与G互相配对的立体化学结果,两条链碱基按照Watson-Crick规则进行配对,其严格性源于A与T,C与G的形状以及氢键的互补特性,需要碱基处于它们优势的异构状态。
双螺旋的稳定性从何而来
- 互补碱基之间的氢键有助于双螺旋的热力学稳定性和碱基配对的特异性。每当一个碱基对形成时在配对碱基之间的产生一个氢键,就有一个碱基与水分子之间的氢键断裂,当双螺旋中的两条链合在一起,水分子被碱基替代,是的双螺旋的以稳定。
- 碱基间的堆积力对于双螺旋的稳定性有重要作用。由于疏水效应,减少了碱基暴露于水分子的可能,从而降低了双螺旋的自由能。
baseflipping
碱基翻出, 单个碱基从双螺旋中突出的现象。碱基从双螺旋中翻出能使碱基处于酶的催化部位,某些酶通过和这些螺旋构型外的碱基相互作用而使碱基甲基化或除去受损的碱基。同源重组和DNA修复相关的酶能将碱基一个接一个地翻转出来,借此扫描DNA搜寻同源区或损伤区域。
DNA通常是右手螺旋
由物理学中的手性法则可知,双螺旋中两条多核苷酸链都是右手螺旋。
实验表明溶液中双螺旋DNA每一螺周含有10.5个碱基对
云母实验表明溶液中双螺旋DNA每一螺周含有10.5个碱基对
- 原理:小片段DNA可以吸附在云母表面上,5’端磷酸基的存在使DNA在云母上成一定的方向性。吸附在云母上的DNA用DNaseI消化,裂解DNA磷酸糖骨架上的磷酸二酯键,但由于酶体积较大,空间位阻使它无法作用于DNA的侧面或底面,仅仅可以切断距离云母最远的DNA表面上的磷酸二酯键,因此切割后得到的片段长度应该可以反映出DNA的周期性,即每一螺周碱基对的数目。在聚丙烯酰胺凝胶电泳中,聚丙烯酰胺凝胶是一种类似果冻的基质,DNA带负电,在胶体中向电场的正极泳动,由于胶体的阻碍作用,片段的泳动速度与片段大小成反比,片段越大泳动速度越慢。
- 实验步骤:吸附在云母上的DNA经过DNase I切割后得到的片段通过聚丙烯酰胺凝胶电泳进行分离。
- 实验结果:从电泳结果中我们发现片段平均大小为 10、11、21、31和 32等,即以 10.5个碱基对逐倍递增的DNA片段成簇存在。每一螺周含有10个碱基对的数值会因为条件不同而有所变动,吸附在云母表面上的DNA每一螺周不止含有10个碱基对。
minor groove & major groove
DNA分子两条链所形成的双螺旋结构使DNA分子延伸成很长的多聚体,其中有两条大小不一的沟,是由碱基对的空间几何结构决定的。碱基对上下堆积,一侧糖间的的窄角形成小沟,另一侧广角形成了大沟。
大沟中有丰富的化学信息
每个碱基对的边缘都暴露于大沟、小沟中,从而产生氢键供体和受体,以及识别碱基对的范德华力表面的模式。
在大沟中的A:T碱基对边缘,化学基团以如下顺序排列:氢键受体、氢键供体、氢键受体和大的疏水表面。ADAM
在大沟中的G:C碱基对边缘,化学基团以如下顺序排列:氢键受体、氢键受体、氢键供体和小的非极性氢。AADH
由于形成的氢键和暴露于大沟中的整体形状的特有模式,可以将A:T和G:C区别开来,也可以将A:T和T:A、G:C和C:G区分开。这些模式使得蛋白质无需解开继而破坏双螺旋就可准确地识别DNA序列。
小沟中化学信息较少,体积小不能容纳氨基酸侧链。
A:T碱基对边缘:氢键受体、非极性氢、氢键受体
G:C碱基对边缘:氢键受体、氢键供体、氢键受体
小沟中不可区分G:C和C:G、A:T与T:A碱基对
但可以区分A:T和G:C碱基对。
双螺旋的多重构象
在较高湿度下观察到的B-DNA最接近生理条件下DNA的一般结构:每螺周10个碱基对,宽的大沟和窄的小沟。
在较低湿度条件下观察到的A-DNA结构:每圈11个碱基对,大沟窄且深,小沟宽且浅。
细胞中绝大部分DNA以B型构象存在,但某些DNA-蛋白质复合体中的DNA也可以以A型构象存在,而且A-DNA与以双螺旋形式存在的RNA分子的结构相似。
B-DNA是一种理想结构,与细胞中的DNA略有不同,体现在两个方面:
- 第一,溶液中的DNA分子比B-DNA分子螺旋程度更高,平均每螺周有10.5个碱基对:
- 第二,B型构象是均一的结构,但实际的DNA没有如此规则,DNA的准确构象取决于双螺旋每一位置上碱基对的种类以及相邻碱基对的存在。
DNA有时可形成左手螺旋
在高阳离子浓度下,中和了带负电的磷酸基团,溶液中交替嘌呤残基才会呈现左手螺旋构象,在低盐浓度下会形成典型的右手螺旋构象。左手螺旋在细胞DNA中只占一小部分。
| 螺旋方向 | 每螺旋单位 碱基数 | 每螺周碱基 数 | 大沟 | 小沟 | ||
|---|---|---|---|---|---|---|
| A型 | 右手 | 1 | ~11 | 极狭、很深 | 很宽、浅 | |
| B型 | 右手 | 1 | ~10 | 宽、中等深度 | 狭、中等宽度 | |
| Z型 | 左手 | 2 | 12 | 平坦 | 极狭、很深 |
denature
变性:双螺旋的两条链依靠较弱的力结合在一起,当DNA溶液温度高于生理温度(接近100$^\circ$C)或者pH较高时,互补的两条链就可以分开,这个过程叫变性,DNA双链完全分开的变性过程是可逆的。
引起DNA变性的因素:
- 加热
- 有机溶剂
- 高pH
- 低盐浓度
reassociate OR annealing
复性/退火:在适当条件下,当变性DNA的热溶液缓慢降温,DNA互补链重新聚合,恢复形成天然规则的双螺旋,这一过程称为复性。
复性应用:由于变性DNA分子具有复性的能力,来源不同的两条DNA通过缓慢降温可以形成人工杂交DNA分子,互补的DNA链和RNA链也可以形成杂交分子,这一特性是Southern印迹杂交和DNA芯片分析等技术的基础。
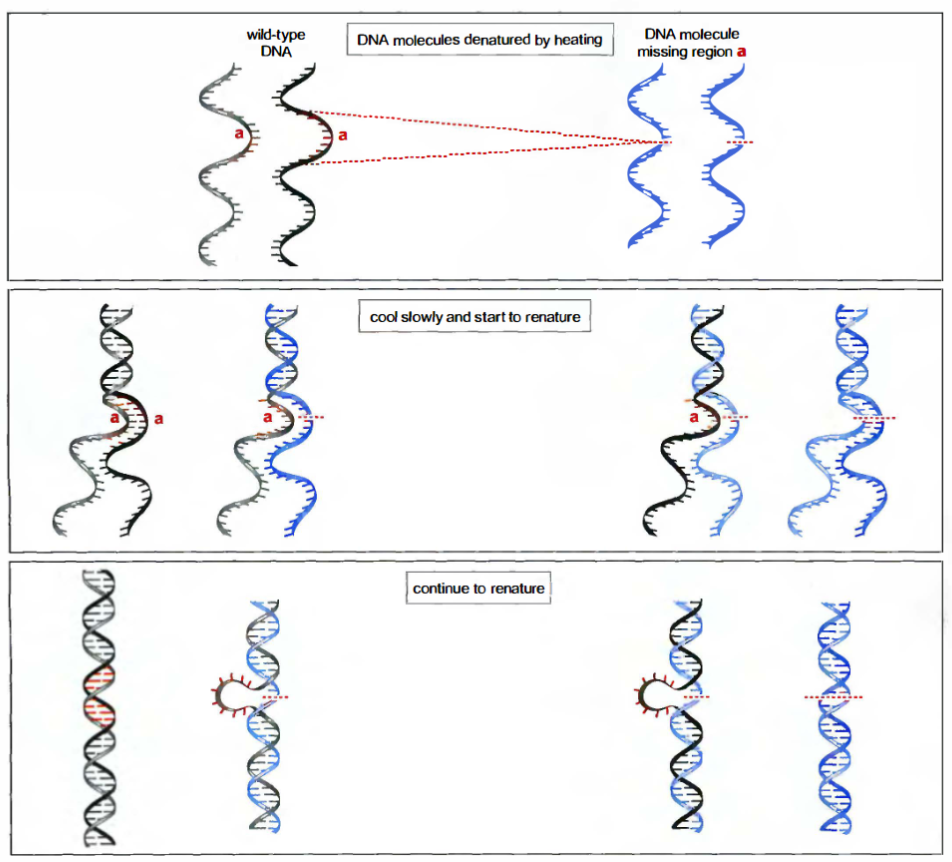
Reannealing & hybridization
退火和杂交。两个基本相同的双链DNA分子的混合物:一个是正常的野生型DNA,另一个是缺少一小段核苷酸的突变体DNA(缺少核苷酸的区域用红色标为a)。加热混合物使DNA变性。变性DNA在低于熔解温度的条件下培育时可以复性,产生两种类型的复性分子:
- 完全复性分子,分别由互补的两条野生型链或两条突变型链构成双螺旋;
- 杂交分子,由一条野生型链和一条突变型链构成,并形成一段短的不配对的环(区域a)。
hyperchromicity
增色效应,DNA在260nm处有最大吸收值,碱基是造成吸收的主要因素。由于双螺旋DNA的光吸收比单链DNA少40%,当DNA溶液温度提高到接近水的沸点时,光吸收值在260nm处明显增加,这一现象称为增色效应。与此相对,减色效应则是因为双螺旋DNA中的碱基堆积降低了紫外线的吸收能力。
melting point
DNA熔点:由DNA的光吸收对温度绘制函数曲线图可知,吸光度增加到最大值一半时的温度叫做DNA熔点,用Tm来表示,Tm值是DNA的特征常数,很大程度上取决于DNA中G+C的百分含量和溶液的离子强度,DNA中的G+C百分含量越高,DNA的Tm值越高,DNA溶液的盐浓度越高,DNA的Tm值也越高。
影响Tm值的因素:
- DNA中G+C的百分含量
- DNA溶液的离子强度
- DNA的均一性:均质DNA的溶解过程发生在一个较小的温度范围内,异质DNA溶解过程发生在一个较宽的温度范围内
- 与DNA分子长度有关,在G+C百分含量差不多的情况下,核酸分子越长,Tm值越大
影响复性效率的因素:
- 温度,比Tm低25$^\circ$C最佳;
- DNA的浓度,浓度越大,复性越快;
- 复性的时间;
- DNA复杂度,高度重复序列DNA复性慢,简单序列比复杂序列快。
DNA Topology
DNA是一种柔性结构,它的精确分子参数与两个因素有关:
- 周围的离子环境;
- 和它形成DNA-蛋白质复合体的DNA结合蛋白的性质。
线性DNA:可以自由旋转以适应双螺旋两条链彼此缠绕数的变化。
环状DNA:如果两条链的糖-磷酸骨架没有断裂,DNA分子两条链彼此缠绕的次数就不能改变,这样的共价闭环DNA(cccDNA)的拓扑结构是受限的。
真核细胞的染色体:长度极长,形成染色质结构,与其他细胞成分相互作用,真核细胞线性DNA分子的拓扑结构也是受限的。
linking number
连环数, 是共价闭环DNA的固有拓扑特性
连环数(linking number, Lk):是指要使两条链完全分开时一条链必须穿过另一条链的次数,连环数是共价闭环DNA的固有拓扑特性,一定是整数。连环数由扭转和缠绕数共同决定。右手螺旋的螺旋交叉数(扭转数)为正数,因此DNA连环数是正值。
连环数由扭转数和缠绕数共同决定
twist:
扭转数(twist, Tw):是一条链旋绕另一条链的螺旋数,即一条链完全缠绕另一条链的次数。cccDNA分子平躺在一个平面上时,这个平面构象中,连环数就等于扭转数。右手螺旋的扭转数定义为正数,因此DNA连环数是正值。
writhe:
缠绕数(writhe, Wr):通常cccDNA不处于一个平面上,而是扭转的,在三维空间里双螺旋的长轴经常重复地自我交叉,这称为缠绕数,缠绕可能有两种形式:互卷和螺线管状。缠绕数是cccDNA中互卷和螺旋管状缠绕数的总和。
Lk$^\circ$是生理条件下完全松驰态cccDNA的连环数
设想cccDNA未形成超螺旋,则它的扭转数就与生理条件下DNA溶液中B型DNA相同,生理条件下cccDNA的连环数(Lk)表示为Lk$^\circ$,这样分子的Lk$^\circ$等于碱基对数目除以10.5所得的值。
这些拓扑学的特性对于理解DNA在细胞内的包装、复制、转录和修复过程中的行为至关重要。DNA的拓扑结构变化,如超螺旋的增加或减少,会对其功能产生显著影响,并且是许多DNA代谢过程的基础。
细胞中的DNA呈负超螺旋
linking difference
连环差(linking difference):用Lk与Lk°的差值表示超螺旋的程度
[ \Delta Lk = Lk - Lk^{\circ} ]
- ( \Delta Lk < 0 ) 时, DNA产生负超螺旋;
- ( \Delta Lk > 0 ) 时, DNA产生正超螺旋。
superhelical density
超螺旋密度(superhelical density): ( \sigma = \Delta Lk / Lk^{\circ} )
细菌和真菌中提取的环状 DNA分子,通常是负超螺旋, ( \sigma ) 值约为 -0.06。
超螺旋的生物学意义:
- 负超螺旋含有自由能,可以为打开双链提供能量,使 DNA复制和转录这样的解离双链过程可以顺利完成;
- 为正超螺旋提供能量,阻止DNA在高温中发生变性;
- 可以让双螺旋扭转数减少。
唯一含有正超螺旋DNA的生物体是哪种?结合 DNA特性进行解释
某些生活在极端高温环境中的嗜热微生物。正超螺旋DNA过度扭曲,正超螺旋提供额外能量,阻止DNA在高温中发生变性,嗜热微生物 DNA双链打开比一般生物呈负超螺旋的 DNA需要更多的能量,因此能在极端高温环境生存。
真核细胞中核小体引入了正or负超螺旋?
真核细胞中核小体引入了负超螺旋
线性DNA本身不能形成超螺旋结构,通过引入核小体, DNA双螺旋以螺线管方式围绕蛋白质核心缠绕两圈,以左手螺旋方式进行盘绕,等同于负超螺旋。
topoisomerase
拓扑异构酶可以使超螺旋DNA解旋
拓扑异构酶(topoisomerase):能在闭环 DNA分子中改变两条链的环绕次数的酶,它的作用机制是首先切断DNA,让DNA绕过断裂点以后再封闭形成双螺旋或超螺旋DNA。
原核生物中存在引导DNA超螺旋形成的特殊拓扑异构酶
促旋酶:原核生物中拥有一种特殊的引入而不是去除负超螺旋的拓扑异构酶II,DNA促旋酶能使原核生物染色体产生负超螺旋,负超螺旋促进DNA双螺旋的解旋,继而激发包括起始DNA转录和复制在内的许多反应。
topoisomerase功能
- 拓扑异构酶可以催化DNA产生瞬时单链或双链的断裂而改变连环数。
- 拓扑异构酶也可以解链和松弛DNA分子。
- 拓扑异构酶通过蛋白质-DNA的共价连接裂解DNA链或使它们重新连接。
- 拓扑异构酶构成酶桥让DNA片段来往穿梭。
topoisomerase作用机制
拓扑异构酶作用机制(以拓扑异构酶I为例):
- 起始解链,拓扑异构酶与DNA片段形成紧密的复合物,复合物中双链变性,一条DNA链结合于酶的裂隙中,使它靠近酪氨酸活性部位的这条链断裂,产生DNA-酪氨酸中间产物,新断裂的DNA的另一末端也被拓扑异构酶紧密结合。
- 断裂后拓扑异构酶经过一次构象变化,在断裂链中打开一个缺口,并通过自身成桥状连接这个缺口。第二条链穿过那个缺口,结合于蛋白质内部的“圆环状”孔中的DNA结合位点。
- 拓扑异构酶-DNA复合物中发生第二次构象变化,攻击磷酸-酪氨酸键的羟基,使DNA的断端重新连接起来。
- 重新连接后,拓扑异构酶发生第三次构象变化,释放DNA,这一过程中DNA产物与最初的DNA分子基本相同,只是连环数增加一个。
以上机制同样适用于拓扑异构酶II,拓扑异构酶II是二聚体,作用的基本特点是DNA双链的断裂需要拓扑异构酶两个亚基及其活性位点中的酪氨酸残基参与。
拓扑异构酶可以不依赖任何其他蛋白质或高能辅助因子,拓扑异构酶活性部位的酪氨酸残基攻击靶DNA骨架上的磷酸二酯键时,DNA断裂产生一缺口,磷酸-酪氨酸连接保存了磷酸二酯键的能量,来自DNA断端的游离羟基攻击磷酸-酪氨酸键,最终重新形成磷酸二酯键。其中拓扑异构酶II需要ATP水解释放的能量,用于拓扑异构酶-DNA复合物的分子构象的改变,使得DNA链断裂或重新连接。
DNA拓扑异构体可以被电泳分离
DNA拓扑异构体(DNA topoisomer):长度相同而连环数不同的共价闭环DNA分子,可以通过琼脂糖凝胶电泳彼此分开,其分离基础是扭曲越大,cccDNA形状就越致密,越容易在凝胶基质中迁移,一个cccDNA的迁移要比这一环状DNA链的高度超螺旋的拓扑结构体慢得多。
DNA环的拓扑学特性证明了DNA每螺周含有个10.5碱基对的螺旋周期性
原理:DNA不同拓扑结构能通过电泳分离。
实验结论:长度相差21个碱基对的两个DNA环迁移率相同,长度相差只有5个碱基对的两个DNA迁移率却不同,因此得出溶液中DNA每周螺旋含有10.5个碱基对的结论。
__离子可使DNA解旋
EB(溴化乙锭):溴化已锭是一种既大又平而且多环的阳离子,它的平面形状使其能滑入或嵌入到堆积的DNA碱基对平面之间,在紫外光下EB会发荧光,而且嵌入DNA后荧光强度显著增强。
功能:
- 作为核酸染料:溴化乙锭因其荧光特性常用于染色核酸,特别是在凝胶电泳后用于观察DNA的存在和迁移模式。
- 可以松弛超螺旋,在一定浓度下,使得负超螺旋变得无超螺旋,如果加入过量,又变为正超螺旋;
- 作为嵌入剂,引起DNA损伤:溴化乙锭嵌入DNA后,可能会干扰DNA的正常结构和功能,导致DNA损伤。
溴化乙锭的使用在分子生物学实验中非常普遍,尤其是在DNA的可视化和分析方面。然而,由于它可能对DNA造成损伤,因此在处理和使用时需要谨慎,并采取适当的安全措施。
名词解释
hybridization
由于变性的DNA分子具有复性的能力,因此来源不同的两条DNA链通过缓慢降温也可形成人工杂交DNA分子,同样互补的DNA和RNA链也可形成杂交分子。两条单链核酸形成杂交分子的能力被称为杂交。是分子生物学中几项不可缺少的技术的基础。
covalent closed circular DNA
共价闭环DNA(cccDNA):游离于染色体之外的质粒双链闭合环形DNA。DNA分子一端的5’磷酸基团与另一端的3’磷酸基团通过磷酸二酯键连接形成的环状DNA分子,是DNA分子的一种特殊形式.
superhelix
超螺旋是指双螺旋DNA进一步扭曲盘绕所形成的特定空间结构,是DNA高级结构的主要形式,可分为正超螺旋与负超螺旋两大类。正超螺旋是指DNA朝着其螺旋方向绕其轴心旋转而成的结构,是过度缠绕的双螺旋。负超螺旋是指DNA朝着其螺旋相反方向绕着其轴心旋转而成的结构。
大题
DNA和RNA之间的结构差异是什么?1.1 DNA和RNA的结构特性如何决定其独特的生物学功能?
- 首先,RNA的骨架含有核糖而不是2’-脱氧核糖。核糖在2’位置有一个羟基。
- 其次,RNA含有嘧啶尿嘧啶而不是胸腺嘧啶。
- 第三,RNA通常是单链多核苷酸,没有互补链。
DNA双螺旋结构模型
DNA的双螺旋结构是沃森和克里克于1953年提出的,DNA双螺旋结构是DNA二级结构的最基本形式。该模型的建立对促进分子生物学及分子遗传学的发展具有划时代的意义。对DNA本身的复制机制,遗传信息的存储形式和遗传信息的表达,生物遗传稳定性和变异等规律的阐明起了非常重要的作用。其主要内容如下:
- DNA分子由两条多核苷酸链组成,两条反向平行的多核苷酸围绕同一中心轴相互缠绕,两条链都是右手螺旋。
- 脱氧核糖和磷酸交替连接,排列在双螺旋外侧,彼此通过3’-5’磷酸二酯键连接,构成DNA分子的基本骨架,碱基对排列在双螺旋的内侧,碱基平面与纵轴垂直。
- 双螺旋的平均直径为2.0nm,相邻碱基平面之间垂直距离为0.34nm,每10个碱基对旋转一圈,碱基对之间的螺距为3.4nm。
- 在双螺旋的表面分别形成大沟与小沟。
- 两条链借助碱基之间的氢键和碱基堆积力牢固结合,维持DNA结构的稳定性。
鉴定DNA还是RNA的方法
加入不含有RNase的DNase1,如果被降解就是DNA,不被降解是RNA。
用二苯胺方法进行测定,如有颜色反应就是DNA,没有就是RNA。
鉴定是单链还是双链的方法
- 可以测定不同碱基所占的比率,如果将合A、T比率相同,G、C比率相同就是双链,如果不符合就是单链。
- 也可以用加热前后260nm处吸光值是否增加来看,如果加热到接近100℃后吸收值高于加热前,就是双链,没有明显增加的就是单链。
DNA比RNA稳定原因
- RNA的磷酸酯键容易被碱水解,而DNA的磷酸酯键不易水解。
- DNA是双螺旋结构,两条链之间存在氢键作用力,且碱基对平面之间具有堆积力,维持DNA稳定性。
- 双螺旋结构相邻碱基对之间的距离为0.34nm,而嘌呤环和嘧啶环的范德华力半径刚好为0.17nm,因此范德华力加强疏水作用。
- RNA的2’-OH比DNA的2’-H更加活跃,故更不稳定。
考点补充
DNA的结构特点
试从DNA和RNA的结构的不同来解释为什么DNA被广泛作为遗传信息的承载者?RNA 在细胞内行使那哪些生物学功能?
- 1.作为遗传物质所需具备的条件
- 1.1.在细胞生长繁殖过程中能够精确地复制自己
- 1.2.能够指导蛋白质合成从而控制生物的性状和新陈代谢
- 1.3.具有储存巨大数量遗传信息的潜在能力
- 1.4.结构比较稳定,但是在特殊条件下又能发生突变,而且突变可以继续复制并遗传给后代。
- 2.DNA比RNA更为广泛地作为遗传信息的承载者的原因(前三点为结构方面原因)
- 2.1.DNA的半保留复制精确地将遗传信息传递给子链而RNA只有一条单链,复制出来的也只是其互补链。而且DNA在复制过程中有修复系统保证其复制的精确性和准确性。
- 2.2.DNA的双螺旋结构比RNA的单链结构更如稳定, 具体体现在:
- RNA的磷酸酯键容易被碱水解,而DNA的磷酸酯键不易水解。
- DNA是双螺旋结构,两条链之间存在氢键作用力,且碱基对平面之间具有堆积力,维持DNA稳定性。
- 双螺旋结构相邻碱基对之间的距离为0.34nm,而嘌呤环和嘧啶环的范德华力半径刚好为0.17nm,因此范德华力加强疏水作用。
- RNA的2’-OH比DNA的2’-H更加活跃,故更不稳定。
- 2.3.在DNA中存在甲基化修饰。甲基化与DNA复制起始及错误修正时的定位有关,还通过改变基因的表达参与细胞的生长、发育过程及染色体中迹、X染色体失活等的调控。甲基的引入不利于模板与RNA聚合酶的结合,从而降低其体外转录活性。有研究表明,甲基化能够很容易地通过翻转碱基和形成Hoogsteen碱基对加以消减,保证DNA双螺旋结挺的稳定性。
- 2.4.DNA聚合酶作用的保真性和校正功能比RNA聚合酶强DNA聚合酶在合成DNA时,其3-5外切酶活性会切割错误掺入的核苷酸,从而修复DNA错配。
- 2.5.DNA损伤修复的机制更完善,确保DNA不发生错误、损伤或者突变
- 错配修复系统: 由错配修复系统识别双螺旋形状上的扭曲,这种扭曲引发一连串的事件,导致含有损伤的一小段单链片段(或小区)被去除。去除修复在 DNA 上产生的单链缺口由 DNA 聚合酶用未受损的链为模板进行填补,恢复原始的核苷酸序列
- 修复酶简单地将损伤逆, 例如光激活作用 (photoreactivation): 光激活作用直接逆转紫外辐射造成的嘧啶二聚体结构。光激活反应中,DNA 光解酶从光线中捕获能量并将其用于断裂连接毗邻嘧啶的共价键
- 碱基切除修复 (base excision repair): 通过修复系统将己变化的碱基除去并替换掉,碱基切除修复作用机制:在碱基切除修复中,一种称为糖基化酶 (glycosylate) 的酶识别并通过水解糖苷键除去受损碱基。在随后的核酸内切步骤中,产生的脱碱基戊糖从 DNA 骨架上去除
- 核苷酸切除修复 (Nucleotide excision repaire, NER): NER主要修复那些影响区域性的染色体结构的 DNA 损害,包括由紫外线所导致的双嘧啶键结,化学分子或蛋白质与 DNA 间的键结,或者 DNA与 DNA 的键结等。蛋白复合体结合到损伤位点,在错配位点上下游几个碱基的位置上将 DNA 链切开,两个切口间的寡核苷酸序列清除, DNA 聚合酶合成新的片段填补间隙,连接酶将新合成片段与原 DNA 链连接起来
- 重组修复 (recombinational repair): 也叫复制后修复,它发生在复制之后。遗传信息有缺损的子代 DNA 分子或者双链断裂的 DNA 可通过重组修复从而加以弥补,即从同源 DNA 的母链上将相应的核苷酸序列片段移至子链缺口处,然后再用新合成的序列来补上母链的空缺,此过程称为重组修复
- 非同源末端连接 (logous end-joining, MEJ): 当DNA双链断裂时,游离的DNA末端吸引二聚体蛋白Ku结合,保护DNA末端防止降解。Ku招募DNA-PKcs及其他蛋白质组装成完整的DNA-PK复合体。DNA-PK复合体促进联会(synapsis)或微同源区 (Regions of microhomology)排列。两个DNA-PK复合体彼此进行磷酸化修饰激活了Ku自身的DNA解旋酶活性。微同源区碱基互补配对,核酸酶降解侧翼(flap), DNA聚合酶填补空隙, DNA连接酶连接切口。
DNA拓扑结构
从大肠杆菌中通过质粒提纯试剂盒提取的质粒在溴乙锭-琼脂糖凝胶电泳上显示为三条带。请问这三条带各代表DNA的哪三种拓扑学结构?设计实验来验证你的推断。
(1) 在溴化乙锭染色的琼脂糖凝胶电泳中,从大肠杆菌中提取的质粒DNA如果显示出三条带,这三条带常代表了DNA的三种不同的拓扑结构:
- 超螺旋(Supercoiled)DNA:这条带通常位于最前方,迁移速度最快。这是因为超螺旋DNA有较高的密度和紧密的结构,使其在凝胶中遇到的阻力最小,迁移速度最快。
- 线性(Linear)DNA:这条带位于中间位置,迁移速度居中。
- 开环(Relaxed)或松弛型(Open Circular)DNA:这条带移动最慢,开环DNA由于其松散的结构,形态像超螺旋DNA那样紧密,所以在凝胶中移动最慢。
(2) 验证实验设计
为了验证这三条带确实代表了DNA的三种不同拓扑结构,可以设计以下实验:
- 1.将原由中的三条带剪下来,胶回收DNA,然后对DNA分别进行下述处理:
- a.酶切:使用适当的限制性核酸内切酶,将超螺旋DNA切成线性DNA。
- b.热变性:通过加热处理,使开环DNA发生热变性,转化为线性DNA。
- c.无处理:将线性DNA直接作为对照。
- 2.重新上样电泳。对经过处理和未处理的DNA样本进行琼脂糖凝胶电泳,观察带型变化。
(3) 实验结果预测:
如果超螺旋DNA经过拓扑结构处理后,原本的超螺旋带消失,出现与原开环DNA相同的带型,则证明超螺旋带是超螺旋DNA。
如果超螺旋DNA样品经过限制性内切酶切割后,出现一条新的带,且位置与原线性DNA相同,则说明超螺旋带是超螺旋DNA。
对照实验:使用未经过处理的质粒标本作为对照,与酶处理的样本进行比较,确认带型的变化确实是由酶处理引起的,从而证明质粒电泳三条带代表的是不同的DNA拓扑结构。
名词补充
Ethidium
溴化乙锭(Ethidium)是一种既大又平而且多环的阳离子,它的平面形状使其能滑人或嵌入到堆积的 DNA 碱基对平面之间从而使 DNA 发生解旋。因为在紫外光下溴化乙锭会发荧光,而且嵌入 DNA 后荧光强度显著增强,因此,溴化乙锭常用来作为染料检测 DNA。
作业
问: 唯一含有正超螺旋 DNA 的生物体是哪种?结合 DNA 特性进行解释。
问: 拓扑异构酶1的作用机制